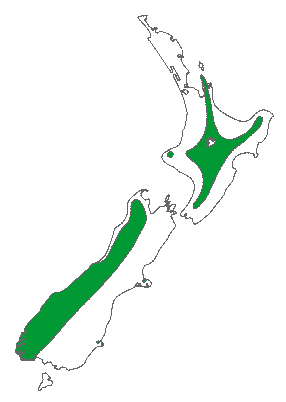
Distribution map (Metcalf 2002).

Mature open-grown tree NW of Mt. Ruapehu [C.J. Earle, 2003.03.08].

Epiphytes and bark on a large tree, Hauhungatahi Track, Mt. Ruapehu area [C.J. Earle, 2003.03.09].

Adult foliage, 4 cm longest dimension [C.J. Earle, 2003.03.09].

Sapling on the Hauhungatahi Track, about 1.5 m tall [C.J. Earle, 2003.03.09].

Juvenile foliage from an understory sapling [C.J. Earle, 2003.03.09].
 .
.
Foliage and cones [Trevor Hinchliffe].

A large tree on the Hauhungatahi Track, 105.5 cm DBH [C.J. Earle, 2003.03.09].

Pygmy forest on the Hauhungatahi Track [C.J. Earle, 2003.03.09].

The pygmy forest - Libocedrus bidwillii about 7 m tall near the brushline on Hauhungatahi [C.J. Earle, 2003.03.09].

Libocedrus bidwillii
Hook. f. 1864
Common names
Pahautea, kaikawaka, mountain cedar.
Taxonomic notes
This is one of those plants that has been recognized as a good, simple species ever since it was first described. There are no synonyms, subspecies or varieties.
Description
Trees to 21 m tall and 100 cm dbh; young plants fastigate, maturing to a long, bare bole with a pyramidal to conical crown of heavy, almost horizontally spreading branches that form distinctively billowing masses. Bark papery, shed in narrow, thin strips. Branchlets of juveniles are somewhat flattened and up to 3 mm broad, the leaves of the two lateral rows closely set. Branchlets of mature plants are tetragonous, bearing four rows of closely set leaves. Juvenile facial leaves are about 1 mm long and the lateral leaves about 3 mm long; adult leaves are subequal, about 2 mm long, triangular, acute and appressed. There are separate male and female cones, which occur on the same tree. Pollen cones are about 7-11 mm long, borne singly at the tips of short branchlets. Seed cones are ovoid, about 7-8 mm long, and are composed of four woody scales, each bearing a curved dorsal mucro. Seeds 2 (one to each fertile scale). Wood is red, soft and straight-grained, but splitting easily (Allan 1961, Dallimore et al. 1967, Salmon 1996, Metcalf 2002).
"Mountain cedar is a handsome and very distinctive tree that can be recognised even from some distance" (Metcalf 2002). It resembles L. plumosa, from which it differs chiefly in its smaller size, smaller leaves and cones, and in the four-sided character of the ultimate branchlets. It is reduced to a bush at higher elevations, or in wet ground (Dallimore et al. 1967).
Distribution and Ecology
New Zealand: N and S Is, at 250 to 1,850 m elevation, in wet forests from latitude 36° 50' southwards. Type locality in the Nelson Mountains (1,830 m) (Allan 1961, Dallimore et al. 1967, Salmon 1996). Hardy to Zone 8 (cold hardiness limit between -12.1°C and -6.7°C) (Bannister and Neuner 2001).
Remarkable Specimens
A specimen 125 cm dbh and 20 m tall is recorded from South Westland, New Zealand (Burstall and Sale 1984).
Dunwiddie (1979), in an exploratory dendrochronological investigation, found a specimen covering 720 years.
Ethnobotany
See Conifers of New Zealand for a review of the historic role of forests in native and European cultures of New Zealand.
The wood is "soft, brittle, durable, less generally useful than that of L. plumosa, but would be suitable for many kinds of carpentry" (Dallimore et al. 1967).
The species has been used in dendrochronology, and is treated in reviews of regional results (Norton & Palmer 1992, Palmer & Ogden 1992, and Xiong and Palmer 2000).
Observations
Salmon (1996) says that a fine, almost pure stand occurs in Hihitahi State Forest east of Waiouru Military Camp, southeast of Mount Ruapehu. As the photographs show, some fine examples can also be found on Mt. Taranaki and Mt. Ruapehu. The finest stand I have seen is along the track up Hauhungatahi, a subsidiary cone west of Mount Ruapehu. To reach this stand, drive 5.5 km south from the highway junction in National Park to the tiny hamlet of Erua and park near the railroad tracks. Follow the tracks about 250 m south to where a small, weathered sign says 'Track' and follow it. The track heads almost continuously uphill, starting in a flax thicket and then, in the forest, passing progressively through elevation zones dominated by rimu (Dacrydium cupressinum), matai (Prumnopitys taxifolia), montane totara (Podocarpus laestus), kaikawaka (Libocedrus bidwillii), yellow pine (Halocarpus biformis), mountain toatoa (Phyllocladus alpinus), bog pine (Halocarpus bidwillii) and pygmy pine (Lepidothamnus laxifolius). The highest subalpine forest has a dense forest of mature Libocedrus bidwillii and Halocarpus biformis only 5-7 m tall, with juvenile plants in the understory. The ecology of the area has been described by Druitt et al. (1990).
Remarks
The epithet bidwillii honors John Carne Bidwill (1815-1853), an English-born Australian botanist who became the first director of the Royal Botanic Gardens, Sydney. Bidwill is also remembered in the names Araucaria bidwillii (Araucariaceae, Australia) and Halocarpus bidwillii (Cupressaceae, New Zealand), and is the only botanist to be honored in the names of so many conifers.
Citations
Allan, H. H. 1961. Flora of New Zealand. Volume I, Indigenous Tracheophyta. Wellington: R.E. Owen Government Printer.
Druitt, D. G., N. J. Enright, and J. Ogden. 1990. Altitudinal zonation in the mountain forests of Mt Hauhungatahi, North Island, New Zealand. Journal of Biogeography 17(2): 205-220.
Xiong, L. and J. G. Palmer. 2000. Libocedrus bidwillii tree-ring chronologies in New Zealand. Tree-Ring Bulletin 56:1-16. See also chronology data at http://www.ncdc.noaa.gov/paleo/metadata/noaa-tree-5365.html, accessed 2007.11.01.
See also
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library. This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
Farjon (2005) provides a detailed account, with illustrations.
Haase, Peter. 1986. A study of a Libocedrus bidwillii population at
Pegleg Flat, Arthur's Pass, New Zealand. New Zealand Journal of Ecology 9:153-156.
Gymnosperms of New Zealand.
The New Zealand Plant Conservation Network, accessed 2010.11.22.





 .
.


