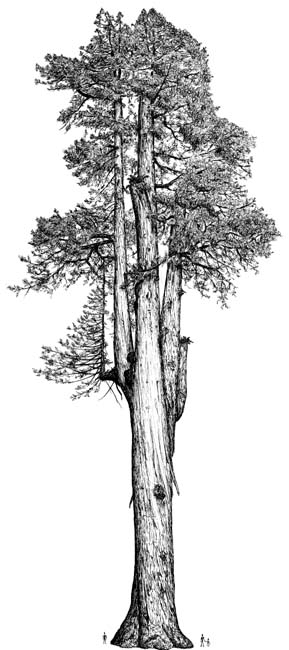
Sequoia sempervirens
(D. Don) Endlicher 1847
Common names
Coast redwood, redwood, California redwood, coastal sequoia, palo colorado; 紅杉 [Chinese]; Séquoia à feuilles d'if [French]; Küstenmammutbaum [German]; セコイアメスギ [Japanese]; 세쿼이아 [Korean]; Секвойя [Russian]; secuoya roja [Spanish]; hồng sam [Vietnamese].
Taxonomic notes
Syn: Taxodium sempervirens D. Don in Lambert 1824 (Watson 1993); Sequoia sempervirens (Lambert) Endlicher (Peattie 1950). It is the sole species in Sequoia Endlicher 1847. Described varieties, all horticultural, include adpressa, glauca, nana pendula, pendula and prostrata (Silba 1986).
Redwood is the only known naturally occurring hexaploid conifer. See Ahuja (2009) for a review of its genetics. Hexaploidy likely arose in the Cretaceous through interspecific hybridization, though there are other possibilities (Ahuja and Neale 2002). The Save the Redwoods League (2017) has recently initiated an effort to fully sequence the Sequoia sempervirens genome.
Description
Tree to 60–100(–110) m tall and 300–460(–900) cm dbh. Trunk much enlarged and buttressed at the base and often with rounded swellings or burls, slightly tapering. Crown crown conic and monopodial when young, narrowed conic in age, irregular and open. Bark red-brown, to ca. 35 cm thick, tough and fibrous, deeply furrowed into broad, scaly ridges; inner bark cinnamon-brown. Branches downward sweeping to slightly ascending. Twigs slender, dark green, forking in a plane, ending in a scaly bud. Leaves 1–30 mm, generally with stomates on both surfaces, the free portion to 30 mm, those on leaders, ascending branchlets, and fertile shoots divergent to strongly appressed, short-lanceolate to deltate, those on horizontally spreading to drooping branchlets mostly linear to linear-lanceolate, divergent and in 2 ranks, with 2 prominent, white abaxial stomatal bands. Pollen cones nearly globose to ovoid, 2–5 mm, borne singly on short terminal or axillary stalks. Female cones 12–35 mm long, elliptical, reddish-brown, with many flat, short-pointed scales; pendant at end of leafy twig; maturing in one season; with 2–5 seeds per scale, light brown, 2-winged. Seeds flattened, 3–6 mm, leathery. 2n= 66 (Little 1980, Watson 1993).
The genus Sequoia has been described as follows: "Trees giant, evergreen. Branchlets terete, with obvious annual growth constrictions. Leaves alternate, mostly in 2 ranks. Adult leaves linear or linear-lanceolate to deltate, generally flattened, divergent to strongly appressed; abaxial glands absent. Pollen cones with 6-12 sporophylls, each sporophyll with 2–6 pollen sacs. Seed cones maturing and opening in 1 season, oblong to globose; scales persistent, 15–30, valvate, ± peltate, thick and woody. Seeds 2–7 per scale, lenticular, narrowly 2-winged; cotyledons 2(–4). x = 11" (Watson 1993).
Distribution and Ecology
USA: SW Oregon and NW California, confined to coastal areas (within 60 km of the sea) experiencing a great deal of fog; at elevations generally below 300 m, occasionally to 1000 m. Mostly found in alluvial soils, where it forms pure stands or occurs with Pseudotsuga menziesii, Chamaecyparis lawsoniana, or other local conifers (Watson 1993). Hardy to Zone 8 (cold hardiness limit between -12.1°C and -6.7°C) (Bannister and Neuner 2001). Sequoia is also very widely planted as an ornamental, found in most areas of suitable climate worldwide, and is locally used in agroforestry, e.g. in New Zealand. See also Thompson et al. (1999).
Distribution data from USGS (1999). Points plotted as tree icons represent isolated or approximate locations.
The "endangered" status assigned this species by the IUCN reflects its past history of exploitation and resulting reductions in its area of occupancy. Recent detailed reviews by Robert Van Pelt (email 2022.09.22) indicate a total area of occupancy of 6,235 km2 within native habitat, which is 25-50% less than other recent estimates. Most conifers with the IUCN "endangered" status have a far more uncertain future than Sequoia. Nonetheless, it is fair to say that in the absence of continuing legal protections afforded the species within its native range, it would be at higher risk than it is now.
Canopy ecology of the redwoods has received a lot of attention in recent years; an excellent summary of the state of knowledge is provided by Sillett and Van Pelt (2007), who document the existence of an essentially complete forest ecosystem (including water sources and storage, nutrient sources and cycling, soil development, and fairly complex animal and plant communities) 60 m off the ground, with a relatively depauperate zone between that height and the ground. This is largely a consequence of trunk reiteration, by which redwoods substitute the usual single-stem architecture of conifers with a multiplicity of stems originating as limbs from a central trunk. In some cases a single tree may have over a hundred such stems, essentially creating a forest from a single tree. Branch surfaces and crotches and rot pockets within this structure provide sites for storage of water and accumulation and development of soils, as well as providing habitat for various mammals, birds, amphibians, and of course a huge arthropod diversity.
On a more terrestrial plane, redwood ecology is well reviewed by Barbour et al. (2001). The subject is really too complex to summarize here, but some important ideas can be discerned from this list of "fun facts":
- On a unit area basis, redwood forests have the highest biomass loadings of any ecosystem on earth.
- Redwoods commonly resprout from stumps or fallen logs (regeneration from seed is almost unknown), which has various consquences. For instance, a redwood forest can be completely destroyed by fire and promptly resprout, a situation which characterizes most redwood stands found on relatively dry sites, such as near the southern and eastern limits of the species' range. Also, redwoods commonly grow in clumps of clonal stems. This means that seedlings may receive carbohydrates, water and nutrition from associated canopy trees, which allows redwoods to outcompete other conifers and to regenerate even in the deep shade beneath their own canopy. It also accounts for the "white redwoods" which have no chlorophyll in their leaves and are entirely supported by the root connections with associated canopy trees.
- Redwood fire ecology includes not only resprout potential, but also fire tolerance mechanisms including highly fire-resistant bark, and an ability to produce epicormic branches in the aftermath of fire. Canopy researchers have found that large trees can lose all live foliage in response to a high severity fire, and then replace it all within the span of a century or so via epicormic branching.
- Redwoods have spectacular height growth, with young trees sometimes growing more than 1 m per year. This places a heavy competitive stress on associated conifers, with the result that the tallest known individuals of Pseudotsuga menziesii and Picea sitchensis are found growing in redwood groves, attaining heights of well over 95 m in their efforts to keep up with the redwoods.
Redwood is one of the few vegetatively reproducing conifers, readily regenerating from stump sprouts in the wake of a major disturbance (typically fire). One consequence of this is the occurrence of 'white' or 'albino' redwoods (see photo), which are trees that originate as root sprouts and are completely nonphotosynthetic, deriving all of their carbohydrate from the roots of their photosynthetic associates (which are not necessarily related, as root grafting is common between redwoods). White redwoods are found only in old-growth forests, where the overstory biomass of photosynthetic redwoods is colossal and the white trees are generally less than 3 meters tall. However, white redwoods up to 20 meters tall are known to exist. The tree in the photograph is about 10 m tall. At the time of the photograph, the new year's foliage had not emerged; trees clad in fresh foliage are snow-white.
Speaking of wildfire, it has been a particularly important issue in Sequoia management during the 21st century. This period has seen the most widespread and severe fires ever observed in native Sequoia forests, with some of the most severe burns incinerating large areas; for example, the 2020 CZU August Lightning Complex Fire affected 35,000 ha, and burned the entire Big Basin Redwoods State Park, producing such widespread and severe damage that the park did not reopen until late in the summer of 2023. I participated in resurvey of an old-growth redwood permanent plot at Big Basin in 2023 and observed that although every redwood in the area had burned, almost no trees larger than 50 cm dbh had been killed, except in the rare cases where a fire had burned a tree so severely that it fell over. Rather, the blackened and charred trunks of the burned trees had all resprouted a new crop of branches, so that they looked like gigantic Italian cypresses (Cupressus sempervirens var. stricta) or, as we called them, "fuzz sticks". To my knowledge, no other tree shows such a tolerance to fire, though it will be interesting to see how long it takes for the fuzz sticks to regain full and complex crown structure. Additionally, virtually all redwoods regardless of size had vigorously stump sprouted, with many of the sprouts exceeding 3 m tall only 3 years after the fire.
Besides wildfire, the other "hot topic" in forest ecology these days is climate change. Redwoods have seen a lot of climate change; during Mesozoic time, the group was widely scattered, and fossils have been found in Asia, Australia, Europe and North America. Forecast climate change is likely to affect extant redwoods primarily at the warm/arid limits of their current distribution (Busiek 2022). However, climate change is also an opportunity for Sequoia, because it potentially leads to introduction and naturalization of these trees at sites widely scattered across the globe, a strategy that could be used to enhance ecosystem resilience as well as to meet societal needs (Antoine and Sillett 2022).
Remarkable Specimens
Big and Tall Trees
The coast redwood is the tallest tree on earth. Older sources speak of extraordinarily tall eucalypts in Western Australia, but thorough searching has documented the heights of those trees in some detail and they are clearly not competitive with coast redwood for height. During historical times there have been Douglas-firs (Pseudotsuga menziesii) recorded as being taller than any coast redwoods now living. However, redwoods growing on benign lowland coastal sites with negligible water stress and very low fire risk were among the earliest victims of the axe and saw, so it reasonable to expect that the tallest trees on Earth were thereby felled during the historical period. We do know that some historically felled redwoods contained larger wood volumes than any tree now living.
The largest volume single-stem tree now living was discovered in Redwood National Park in 2014 by Mario Vaden and Chris Atkins. Based on multiple measurements by highly credible observers including Chris Atkins, Michael Taylor, and Ron Hildebrandt, the tree, which is named Grogan's Fault, has an estimated stem volume of 1,084.5 m3 (38,299 ft3). This is all the more impressive because it is clearly a single-stem tree (Vaden 2015, Mario Vaden email 2015.05.20). A larger volume is found in the Lost Monarch, 1205 m3 and 98 m tall (Sillett [no date]). This is a multiple-stem tree from a single genetic individual (a clone) in which the wood of the separate stems is completely fused to a substantial height.
The Crannell Giant may well have been the largest coast redwood ever cut. Taylor and Mifsud (2010) report that it formerly grew about a mile southeast of Big Lagoon, north of Trinidad, California. This tree carried a total stem volume of at least 1,743 cubic meters and was 94 m tall. A cookie from this tree makes up one wall of the "One Log Tree House" tourist attraction on Broadway in Eureka, California. Logged in about 1945, this was the largest tree ever measured, with a total aboveground wood volume about 18% larger than the largest Sequoiadendron now living. It proved to be extremely rotten, and yielded little merchantable timber.
The following table (source, Michael Taylor email 2009.10.17 with some later updates), provides summary information on all coast redwoods known to be over 110 meters tall, as measured by direct tape drop or laser by surveyors including Steve Sillett, Robert Van Pelt, Chris Atkins, Ron Hildebrant, and Michael Taylor. Most of these trees are remeasured at intervals of no more than three years; the tallest are remeasured yearly. As a point of interest, when I first published this table in 1998, it had only 12 trees. At this point, though, nearly all trees over 110 m have likely been discovered, and none have been found outside of the parks shown below, despite extensive surveys. The inventory of trees 100 to 110 m tall, though, covers much of the species' range, and many new trees are discovered yearly; as of April 2022 the tally of trees over 100 m tall stands at 2,585 individuals, and more are discovered every year. The names in the following table are widely used by people familiar with the largest redwoods, and many of them have appeared in the popular press, such as in Van Pelt (2001) and Preston (2007), as well as in the scientific literature, e.g. Sillett and Van Pelt (2007). The use of such names personifies these trees as individuals and thus helps foster support for their continued protection. However, it may attract undue attention to individual trees, which can lead to their harm. Thus the precise locations of these trees are not presented here, and should remain secret.
| | | | |
Tree | Park | Diameter (m) | Height (m) |
| Hyperion | Redwood National Park | 4.84 | 115.85 |
| Helios | Redwood National Park | 4.96 | 114.58 |
| Icarus | Redwood National Park | 3.78 | 113.14 |
| Stratosphere Giant | Humboldt Redwoods State Park | 5.18 | 113.05 |
| National Geographic | Redwood National Park | 4.39 | 112.71 |
| Orion | Redwood National Park | 4.33 | 112.63 |
| Federation Giant | Humboldt Redwoods State Park | 4.54 | 112.62 |
| Paradox | Humboldt Redwoods State Park | 3.90 | 112.51 |
| Mendocino | Montgomery Woods State Reserve | 4.19 | 112.32 |
| Millennium | Humboldt Redwoods State Park | 2.71 | 111.92 |
| Apex | Humboldt Redwoods State Park | 3.38 | 111.83 |
| Pipe Dream | Humboldt Redwoods State Park | 4.27 | 111.71 |
| Harry Cole | Redwood National Park | 4.94 | 111.65 |
| Rockefeller | Humboldt Redwoods State Park | 4.84 | 111.59 |
| Minaret | Humboldt Redwoods State Park | 3.66 | 111.56 |
| Alice Rhodes | Humboldt Redwoods State Park | 3.35 | 111.53 |
| Mother & Daughter | Humboldt Redwoods State Park | 3.60 | 111.50 |
| Lone Fern | Humboldt Redwoods State Park | 2.56 | 111.47 |
| Teepee Bell | Montgomery Woods State Reserve | 4.08 | 111.34 |
| Paul Zinke | Humboldt Redwoods State Park | 2.90 | 111.31 |
| Aether’s Arrow | Humboldt Redwoods State Park | 3.38 | 111.16 |
| Paul Zahl | Redwood National Park | 3.78 | 111.16 |
| Daedalus | Redwood National Park | 4.26 | 110.77 |
| Rocket Top | Humboldt Redwoods State Park | 3.29 | 110.77 |
| Pinnacle | Humboldt Redwoods State Park | 3.05 | 110.71 |
| Harriett Weaver | Humboldt Redwoods State Park | 4.11 | 110.68 |
| Pyramid Giant | Humboldt Redwoods State Park | 5.03 | 110.68 |
| Valentine | Humboldt Redwoods State Park | 3.47 | 110.65 |
| Libby (A.K.A. Tall) | Redwood National Park | 2.90 | 110.62 |
| Tranquility | Humboldt Redwoods State Park | 4.14 | 110.55 |
| Crown Jewel | Redwood National Park | 3.54 | 110.40 |
| South Fork | Humboldt Redwoods State Park | 4.02 | 110.40 |
| Springing Buck | Humboldt Redwoods State Park | 4.79 | 110.20 |
| Swamp | Montgomery Woods State Reserve | 3.41 | 110.09 |
| John Muir | Humboldt Redwoods State Park | 4.27 | 110.06 |
| T4 | Redwood National Park | 3.99 | 110.06 |
| Rockview | Humboldt Redwoods State Park | 3.84 | 110.03 |
| | | | |
Sequoia is also the tallest horticultural conifer. Several trees in the memorial grove at Whakarewarewa in Rotorua, New Zealand, have been measured by laser or tape drop to be over 70 m tall, including one at 71.2 m (Smillie 2013), and two measured by tape drop at 71.5 and 70.5 m tall (Arborfilm 2014) (however, the tallest tree in New Zealand is a Eucalyptus that exceeds 80 m).
Old Trees
The oldest trees on record exceed 2200 years. I have prepared a report on a section from a log in Humboldt Redwoods State Park that has 2267 rings. The specimen, which has poor circuit uniformity with many missing rings, was scanned and crossdated by Allyson Carroll at Humboldt State University in 2013; the record evidently extends from 385 BC to 1881 (2,266 years), of which the 1071-1813 period has been crossdated with other Sequoia samples. Besides this specimen, a ring-counted age of 2200 years was reported for a specimen from N California collected by Fritz (Brown 1996). This was probably a stump count. There is also said to be a cross-section on display at the visitor center in Henry Cowell State Park, Santa Cruz, CA, that has 1935 rings with the center of the log missing; the original tree was felled in Humboldt County (Kenyon Moon email 2007.11.04).
With many old trees, there arises the question, can they live forever? In principle, yes. There is little evidence supporting the existence of senescence in conifers, and all really old trees (discussion) live many times longer than the oldest living cells in the tree. In practice, though, there are certain environmental factors that can kill trees. When a tree is resistant to those factors, and/or when the factors are expressed weakly or rarely, then the tree may attain a great age. The environmental factors never go away, though, and it is a statistical inevitability that every tree will eventually meet its end. Big redwoods are killed mostly by some combination of fungus, wind, gravity, fire, and flood. The most common death is due to stem or root rots that leave the tree vulnerable to breakage by wind, gravity, fire, and flood. Gravity kills trees when they develop a lean, which never gets better and usually gets worse, eventually breaking or uprooting the tree, often during a big storm when winds stress the crown and flooded soils reduce friction between roots and soil. Fire, if not immediately fatal, can kill by repetition: one fire opens a fire scar through the thick bark, and later fires enlarge the scar and allow fungi to enter the heartwood, so that the tree will eventually break off. Floods usually kill redwoods when streambank erosion undercuts and topples a tree on a floodplain. There are also freak accidents, like when a falling tree hits another tree and delivers an injury that eventually proves fatal. And of course there are chainsaws. None of these things are common (except chainsaws), but 2,000 years is a long time.
A few years ago, dendrochronology suffered a bit of a scandal—or at least, what might pass for a scandal in this peaceful field of study—when a person having no knowledge of the subject represented themselves as an expert to someone at the Guinness Book of World Records, claiming to have found a 12,000 year old redwood tree, which they modestly named the Eternal God Tree. Guinness duly published the claim. Later that year, I was shown the tree and had the opportunity to sample it with an increment borer. The sample produced 515 years and the tree is rotten inside of that. Given the tree's relatively small size (about 3 m in diameter, which, yes, is "small" in comparison with other trees nearby), I would very much doubt it could be a day over 2,000 years old, and my best guess is about 1,200 years—a tenth of its claimed age. Incidentally, the 2,266-year-old tree described above was assigned an age of 7,000 years by the same person. I only mention this because you may see these fanciful ages of 7,000 and 12,000 years reported in the news media.
Popular Culture
Humans have put Sequoia to one use that is not seen in any other tree, except a few Sequoiadendron. We drive through them. Here is a brief history of drive-through trees; all data from Van Pelt (unpublished information transmitted 2022.09). Note that the number of drive-through trees is diminishing; even in such large trees, cutting a tunnel severely reduces the tree's structural strength, and most of the historic tunnel trees have subsequently fallen. No new tunnel trees are likely to be created, at least on public lands.
| | |
Tree | Remarks |
| Coolidge Tree | Tree located near Leggett, California. Tunnel made between 1910 and 1915, named for Col. John Coolidge, father of the President. Area named Underwood Park in the 1920s. This was a relatively small tree and became unstable, and was felled in 1938. |
| Shrine Tree | On private land near Humboldt Redwood State Park. Tunnel was cut by enlarging an existing opening in the hollow tree, perhaps as early as 1920. This short, broken, tree only has a few live branches near the top. It remains open for vehicles. |
| Chandelier Tree | Also in Underwood Park, this is a considerably larger tree with a superb crown. Tunnel was cut in 1937. The tree remains standing and is now the largest tree open to drive through. |
| Tour Thru Tree | Located near where US 101 crosses the Klamath River, this tunnel through a relatively small redwood was cut in 1976. it remains open to vehicles. |
| Drive-Way Stump | This was a tunnel cut through the base of the stump of a logged tree, and was in use in the 1930's-1940's, approximately. It was located at the north end of the Avenue of the Giants, near Pepperwood, California. |
| Twin Drive Thru Tree | This was two smaller redwoods fused through the base, which allowed cutting a tunnel large enough to carry a small (about 12-inch gauge) excursion train. It was located at Confusion Hill, tourist attraction north of Leggett, and seems to have been active in the 1960s-1970s. |
| Smokestack | A tunnel through Smokestack, a relatively small redwood, now conveys the excursion train at Confusion Hill. |
| | |
Ethnobotany
Exploitation and Conservation
There was a time when redwoods were the largest trees on earth, and nearly all were cut in a frenzy of logging activity that lasted over a century, beginning when California was Spanish and not ending until the 1990s, when the last old-growth grove on timber production lands was cut. The film Redwood Saga, made in 1946, provides an excellent illustration of how logging was performed at that time, and this 1895 article (accessed 2023.03.11) discusses the many and varied uses of redwood lumber at that time.
The most severe period of logging began in about 1850, driven by the demand for mine and building timber created by the California gold rush, and continued well into the 20th century. Opposition to the logging began almost immediately; in 1852, Assemblyman Henry A. Crabb of San Joaquin County asked the California legislature to secure into public ownership all of the state's redwood forest lands. Efforts at conservation started to bear fruit in 1900, when the Sempervirens Club was established (Sempervirens Fund [no date]). This was the first activist organization to formally work to preserve the trees, and two years later, the first redwood park (now Big Basin Redwoods State Park) was set aside. Muir Woods National Monument was preserved in 1908, and in 1918 the Save The Redwoods League was established. In subsequent decades the League and many other activist groups (for a detailed review see Speece 2017) fought to protect remaining old redwoods by advocating for state and federal parks, securing donations of land, and sometimes buying land outright. Today, the redwoods are one of the most symbolically important of all tree species, revered by millions of people for the aesthetic and other intangible values provided by these remaining groves of immense, majestic trees.
See also:
- Harper's article, 1883, about a redwood logging camp.
- Harris, D. 1997. The Last Stand: The War Between Wall Street and Main Street over California's Ancient Redwoods. Sierra Club Books.
- Hill, J.B. 2000. The Legacy of Luna. Harper.
- Yaryan, W., D. Verardo and J. Verardo. 2000. The Sempervirens Story: A Century of Preserving California's Ancient Redwood Forest, 1900-2000. Sempervirens Fund Press.
Managed redwood plantations now cover large expanses of private land in northern California, and in them this very fast-growing species produces commercial crops of a lumber valued for its beauty, strength, light weight and decay resistance. It is widely used for outdoor products such as decks, lawn furniture, planters, hot tubs, etc.
Popular Culture
Sequoia is a remarkably popular trademark and marketing theme, that has been applied to (very large) finance companies, (very large) cars, tech and biotech companies, parks, schools, restaurants, beers, wines, and almost countless other commercial products ("almost": Google provides about 33 million hits on "Sequoia company" and 22 million on "Sequoia products"). This is likely a consequence of the tree's extraordinary level of influence on the public imagination and the creative process, a theme explored in detail by Vermaas (2003).
Redwoods have been featured in quite a few movies. Some of the more memorable examples include the Endor chase scene in the Star Wars film Return of the Jedi (1983) and a 1952 drama, The Big Trees, starring Kirk Douglas as a dimpled logger. The scenes in Return of the Jedi were mostly filmed in a stand on Miller-Rellim Redwood Company land that was logged soon after the filming, which is probably why they were able to get permission to film on such a site. Some shorter sequences were filmed at the Owen Cheatham Grove in Grizzly Creek Redwoods State Park, and this site is now quite popular with Star Wars fans (Vaden 2016). On a more scientifically respectable level, redwoods also co-star in parts of the BBC miniseries Walking With Dinosaurs, which also features great photography in such gymnosperm-rich locations as Chile and New Caledonia. These are among the few places left where you can see the world as it looked to the dinosaurs. Since that time the radiation of angiosperms and the appearance of grasses have dramatically remade most of the world's terrestrial ecosystems.
Dendrochronology
Sequoia has proven to be very useful in dendrochronology. Fritz (1940) concluded that the species is not generally useful due to poor circuit uniformity (i.e. the rings do not go all the way around). However, a more exhaustive study by Schulman (1940) found that circuit uniformity improves when samples are taken well above the base of the tree; he successfully crossdated samples from cross-sections taken (from logged trees) 30 m above the ground, and found evidence of narrow rings in drought years. More recent work (Carroll et al. 2014) has confirmed Schulman's findings and used tree-ring width data from a collection of 8 chronologies representing 76 trees, collectively spanning the species' latitudinal distribution. This study only managed to crossdate about 83% of the ring-width record, which is an unusually low fraction, indicating that circuit uniformity and related problems remain a substantial obstacle to dendrochronological use of the species. Nonetheless, the data for crossdated specimens were used study the climate-growth response in this species, and to develop a long reconstruction of climate variability. Increment core samples are limited by the available equipment, rarely delivering a core more than 60 cm long. However, because most of these large trees are 2-3 m in diameter at the base of the live crown, a 60 cm core may record over 1,000 years of record, and several of the chronologies developed by Carroll et al. (2014) have a useful sample depth (i.e., more than about 20 cores) over a period of close to or more than 1,000 years. As with most species, trees near the northern range limit show a growth response strongly correlated with growing season temperature, while trees near the southern range limit have a comparably strong response to growing season precipitation (Carroll et al. 2014).
Observations
The species is well protected in its native range and can easily be seen in California's Redwood National Park, Muir Woods National Monument, and a long chain of State Parks of which some of the more noteworthy are, from north to south, Jedediah Smith Redwoods, Prairie Creek Redwoods, Humboldt Redwoods, Montgomery Woods, and Big Basin Redwoods. If you have never seen a grove of ancient redwoods, you should do so. It is one of the finest sights anywhere on the planet.
Remarks
The genus is widely thought to be named for Sequoyah, also known as George Guess, inventor and publisher of the Cherokee alphabet. Endlicher was also a philologist, so he likely knew of Sequoyah's achievements. Unfortunately, Endlicher's writings give no clue to the etymology of Sequoia, and some very eminent botanists have proposed plausible alternatives. Asa Gray, for instance, thought it came from the Latin sequi, 'following', since it is the sole living representative of a sizable group of extinct plants (Hartesveldt et al. 1975). The epithet sempervirens is Latin for "always green" and was given because the species was originally placed in Taxodium, a genus of deciduous trees.
The sequoias (including Sequoia and Sequoiadendron) were another group, like Metasequoia, first known from the fossil record, although fossil material was not formally named until Steinhauera Presl 1838, ten years after Lambert described this species as Taxodium sempervirens (Hartesveldt et al. 1975). The genus has a rich fossil record in western North America, represented by the Eocene and Oligocene fossil taxon Sequoia affinis and the pollen morphogenera Taxidiaceaepollenites and Sequoiapollenites. Well-preserved examples of S. affinis cones, foliage and wood—including in situ stumps over 200 cm dbh—have been found at Florissant Fossil Beds National Monument (National Park Service 2022), at Copper Basin in Nevada (Inyo [no date]), and at various other locations in the West. These fossils bear a close resemblance to living S. sempervirens. The genus is also represented by the late Miocene Sequoia maguanensis of Yunnan, China, the well-preserved foliage of which bears a strong resemblance to the shade foliage of S. sempervirens, suggesting Asian-North American continuity of habitat across the Bering land bridge at or about that time (Zhang et al. 2015).
Redwoods (Sequoia sempervirens and Sequoiadendron giganteum) are the state tree of California (Watson 1993).
The first detailed description of a sequoia was written by Father Juan Crespi, a friar of the Portola expedition in 1769. Father Pedro Font took the first measurements (girth and height) of a coast redwood in 1776. This tree still stands and gave its name to the city of Palo Alto. In 1791, Thaddeus Haenke, a Czech botanist of the Malespina expedition made the first botanical description of the tree (Meyer 2014). A large part of his
collections was shipped to Spain between 1791 and 1799, and the species was early (ca. 1826) introduced to Spain thanks to the collections of Tadeas Haenke during the Malaspina expedition; an account of these trees appears HERE.
Citations
Ahuja, M.R. 2009. Genetic constitution and diversity in four narrow endemic redwoods from the family Cupressaceae. Euphytica 165:5-19.
Ahuja, M.R. and D.B. Neale. 2002. Origins of polyploidy in coast redwood (Sequoia sempervirens (D. Don) Endl.) and relationship of coast redwood to other genera of Taxodiaceae. Silvae Genetica 51:93-100.
Antoine, Marie and Stephen Sillett. 2022. Intentional forests: growing hope for the future. International Dendrology Society Yearbook 2021, pp. 54-67.
Arborfilm. 2014. The Redwoods of Rotorua - A Tree Climbers Journey. Documentary film on Youtube. Accessed 2016.02.24.
Barbour, M.G., S. Lydon, M. Borchert, M. Popper, V. Whitworth and J. Evarts. 2001. Coast Redwood: A Natural and Cultural History. Los Olivos, CA: Cachuma Press.
Busiek, Julia. 2022. Redwoods and climate https://sempervirens.org/news/redwoods-and-climate/, accessed 2023.11.05.
Carroll, A. L., S. C. Sillett, and R. D. Kramer. 2014. Millennium-Scale Crossdating and Inter-Annual Climate Sensitivities of Standing California Redwoods. PLOS ONE 9(7):e102545.
Fritz, E. 1940. Problems in dating rings of California coast redwood. Tree-Ring Bulletin 6(3):19-21.
Inyo. [no date]. Field Trip To The Copper Basin Fossil Flora, Nevada. http://members.aol.com/Waucoba7/cb/copperbasin.html, accessed 2006.10.03, now defunct.
Meyer, Marc. 2014. When men met the redwoods. European Champion Tree Forum, 4th Meeting, Valsaín, Spain, 10th to 14th October 2014. Ahttp://www.championtrees.eu/reports/ECTF_2014_Report.pdf, accessed 2017.09.03, now defunct.
National Park Service. 2022. Sequoia affinis reconstruction. https://www.nps.gov/flfo/learn/nature/sequoia-affinis.htm, accessed 2023.11.05.
Save the Redwoods League. 2017. Redwood Genome Project. Available https://www.savetheredwoods.org/project/redwood-genome-project/, accessed 2017.10.04.
Schulman, E. 1940. Climatic chronology in some coast redwoods. Tree-Ring Bulletin 6(3):22-23.
Sempervirens Fund. [no date]. History of Sempervirens Fund. http://www.sempervirens.org/history.htm, accessed 2008.08.08, now defunct.
Sillett, S. C. [no date]. Redwood photo tour. http://www.humboldt.edu/~sillett/photos/sese/championtrees/full/3_metric.jpg, accessed 2008.08.07, now defunct.
Smillie, Matt. 2013. Record BPR/1208. https://register.notabletrees.org.nz/tree/view/1208, accessed 2019.03.01.
Speece, D. F. 2017. Defending Giants: The Redwood Wars and the Transformation of American Environmental Politics. Seattle: University of Washington Press.
Taylor, Michael and Brett Mifsud. 2010. http://www.landmarktrees.net/index.html, accessed 2010.10.31, now defunct.
Vaden, Mario. 2010. Largest & Tallest Coast Redwoods, Parks, Photos, accessed 2010.02.12. Lots of interesting information, links, and great photos.
Vaden, Mario. 2015. Largest Coast Redwood ~ 1 (a). http://www.mdvaden.com/redwood_new_era.shtml, accessed 2015.07.26, now defunct.
Vaden, Mario. 2016. Redwoods & Star Wars, Return of the Jedi, Endor, accessed 2020.04.06.
Vermaas, Lori. 2003. Sequoia: The Heralded Tree in American Art and Culture. Smithsonian Books.
Zhang, Jian-Wei, Ashalata D’Rozario, Jonathan M. Adams, Ya Li, Xiao-Qing Liang, Frédéric M. Jacques, Tao Su, and Zhe-Kun Zhou. 2015. Sequoia maguanensis, a new Miocene relative of the coast redwood, Sequoia sempervirens, from China: implications for paleogeography and paleoclimate. American Journal of Botany 102(1):103-118.
See also
Brown, J.E. 1982. Monarchs of the Mist: The Story of Redwood National Park and the Coast Redwoods. Redwood Natural History Association.
Davis, Douglas F. and Dale F. Holderman. 1980. The White Redwoods. Happy Camp, CA: Naturegraph. 45pp.
Del Tredici, Peter. 1998. Lignotuber formation in Sequoia sempervirens: development and ecological significance. Madroño 45: 255-260.
Del Tredici, Peter. 1999. Redwood burls: immortality underground. Arnoldia 59(3): 14-22.
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library. This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
Farjon 2005.
Johnstone, P and P. E. Palmquist. 2001. Giants in the Earth: The California Redwoods. Heyday Books.
Meyer, H.W. 2003. The Fossils of Florissant. New York: HarperCollins.
Moore, Zane. [no date, ca. 2013]. Coast redwood albinism and mosaicism. http://www.mdvaden.com/documents/albino_redwoods_chimera.pdf, accessed 2014.03.29, now defunct. Excellent discussion and bibliography for "white" or "albino" redwoods.
Noss, R.F. (ed.). 1999. The Redwood Forest. Island Press.
O'Hara, K. L. and J.-P. Berrill. 2009. Epicormic sprout development in pruned coast redwood: pruning severity, genotype, and sprouting characteristics. Annals of Forest Science 66:409-417.
Schwarz and Weide (1962).
Threatened Conifers of the World.
Vaden, Mario. Coast Redwood Adventures. http://www.mdvaden.com/grove_of_titans.shtml, accessed 2019.02.24. Lots of detail on the largest trees, and lots of outstanding photos.
Wingate, F.H. and D.J. Nichols. 2001. Palynology of the Uppermost Eocene Lacustrine Deposits at the Florissant Fossil Beds National Monument, Colorado. In: Fossil Flora and Stratigraphy of the Florissant Formation, Colorado; Proceedings of Denver Museum of Natural Science: Series 4, No. 1.