Pinus monticola
Douglas ex D. Don in Lambert 1832
Common names
Western white pine; silver, soft, fingercone, mountain, Idaho, or little sugar pine (Peattie 1950); pin argenté (Kral 1993).
Taxonomic notes
Syn: Strobus monticola (Douglas ex D. Don) Rydberg (Kral 1993). One of 23 species in Pinus sect. Quinquefoliae. The most comprehensive analysis of Pinus yet performed found P. monticola sister to a clade containing the species in the P. flexilis-P. ayacahuite clade, nested within a clade that also included P. strobus, P. chiapensis, and P. lambertiana (Jin et al. 2021), and this result agrees well with past molecular studies (Liston et al. 2007, Syring et al. 2007) as well as with traditional morphologically based interpretations (e.g., Price et al. 1998).
Description
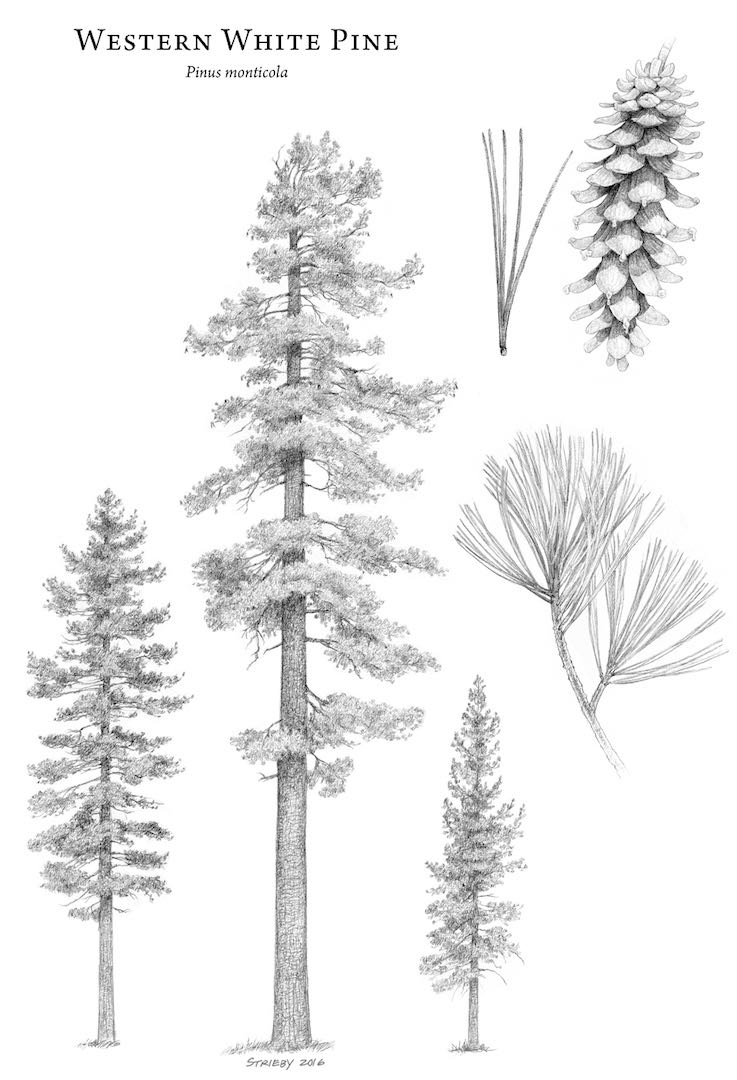
Trees 30(70) m tall and 100(250) cm in diameter, straight; crown narrowly conic, becoming broad and flattened. Bark grey and thin, smooth, becoming furrowed into distinctive rectangular to hexagonal scaly plates in large individuals. Branches nearly whorled, spreading-ascending; twigs slender, pale red-brown, rusty puberulent and slightly glandular (rarely glabrous), aging purple-brown or gray, smooth. Buds ellipsoid or cylindric, rust-colored, 0.4-0.5cm, slightly resinous. Needles 5 per fascicle, spreading to ascending, persisting 3-4 years, 4-10 cm x 0.7-1 mm, straight, slightly twisted, pliant, blue-green, abaxial surface without evident stomatal lines, adaxial surfaces with evident stomatal lines, margins finely serrulate, apex broadly to narrowly acute; sheath 1-1.5 cm, shed early. Staminate cones ellipsoid, 10-15 mm, yellow. Ovulate cones maturing in 2 years, shedding seeds and falling soon thereafter, clustered, pendent, symmetric, lance-cylindric to ellipsoid-cylindric before opening, broadly lanceoloid to ellipsoid-cylindric when open, 10-25 cm, creamy brown to yellowish, without purple or gray tints, resinous, stalks to 2 cm; umbo terminal, depressed. Seeds compressed, broadly obovoid-deltoid; body 5-7 mm, red-brown; wing 2-2.5 cm. 2n=24 (Kral 1993, Little 1980). The most conspicuous features of this species are the hexagonally-checked bark of large trees, unlike that of any other tree in North America; and the exceptionally large cones, longer than those of any other pine in its range except in California, where they resemble miniature Pinus lambertiana cones. Young trees of the two species can be distinguished by the slightly darker, thicker, stiffer needles of P. lambertiana, and the needles of P. monticola are minutely serrate (easier to feel than to see), which distinguishes it from the smooth-edged needles of P. albicaulis.
Distribution and Ecology
Western USA: Washington, Montana, Idaho, Nevada, Oregon, California; and Canada: Alberta, British Columbia; elevations to 1000 m in N, and at 1900-3000 m in the S. Occurs in lowland fog forests or on moist mountain soils, occasionally in forested bogs. Usually in mixed conifer forests, occasionally in pure stands (Kral 1993, Little 1980). See also Thompson et al. (1999). Hardy to Zone 4 (cold hardiness limit between -34.3°C and -28.9°C) (Bannister and Neuner 2001, but provenance unknown; see remarks below).
Ecologically, P. monticola is most often found as a secondary component of mixed-conifer stands, commonly in association with relatively mesic, shade-tolerant trees such as Abies and Tsuga, although not infrequently with Pinus or Pseudotsuga. It is best adapted to two quite distinct environments. In one, it does best on good soils with adequate water, where it can outgrow most xeric-adapted species (such as Pinus subsp. Pinus, or Pseudotsuga) but suffers from competition with shade-tolerant species (such as Abies and Tsuga) that nonetheless are intolerant of fire; in these settings, low-intensity ground fire favors P. monticola over the shade-tolerant species, and so it was historically most prolific in areas having such a fire regime. The second environment where it does well is on extremely poor soils, such as bogs, sands, glacial outwash gravels, and barren rock. In these places it is second only to P. contorta in its ability to survive, and sometimes even to prosper.
Distribution based on herbarium collections and iNaturalist observations; open left map panel for details.
A common garden experiment using quantitative traits and genetic analysis linked to a large selection of climate variables was used by Richardson et al. (2009) to show that there are two major climate response ecotypes in P. monticola, found in trees respectively north and south of an abrupt transition zone in the southern Oregon Cascades. Trees south of this line experience warmer, drier summers, have lower tolerance for low temperatures, and have lower growth potential; there is also a gradient in cold tolerance from maritime to continental sites. This amounts to a scientific confirmation of something that has long been noted, a clear distinction between the white pines of the Pacific Northwest and those of California (including the Warner, Klamath, and Siskiyou Mountains, and the Sierra Nevada). In the northwest, the species' ecology is largely as described above, but in the Sierra, it becomes a species of the high mountains, sharing its habitat with other conifers of California's mixed subalpine cover type, including Abies magnifica, Tsuga mertensiana, Pinus contorta subsp. murrayana, Pinus albicaulis, and Pinus balfouriana; and at lower elevations on barren sites such as granite domes, where it occurs with species such as Pinus jeffreyi and Juniperus grandis. As such it is a generally more xeric habitat than in the northern populations, with lower biomass loadings and lower plant species diversity. It is also a habitat largely without fire, likely due to open forest structures and low fuel loadings.
Remarkable Specimens
The official largest tree is a pine on the California/Nevada border (slightly on the Nevada side), near Lake Tahoe. It measures 263 cm DBH and is 39 m tall (American Forests 2019, Bob Van Pelt pers. comm. 2019.07.13). Formerly, all the really big white pines grew in Idaho, but they have been largely destroyed by logging and white pine blister rust (Ethnobotany, below). For instance, Peattie (1950) reports harvest of trees 175 feet tall and 8 feet dbh (53.3 m and 244 cm, respectively). The remaining giant Idaho pines are in the Floodwood State Forest, where you can find the Floodwood Giant (52 m3 stem volume, dbh 201 cm, height 69.2 meters) and the tallest known white pine, reaching 70.7 meters high (Van Pelt 2000).
Harlow and Harrar (1969) report a maximum age of 615 years. The source is not stated, but most of their measurements are from ring counts on stumps. Some of the Sierra Nevada trees appear older, but I have found no data.
Ethnobotany
Aboriginal uses were both material and medicinal. Material uses focused on the pitch, which was used to coat whaling and fishing equipment, and to fasten arrowheads to shafts (Arno and Hammerly 2007). Medicinal uses mainly involved boiling the bark and drinking the infusion. The Quinault used this for disorders of the stomach and to purify the blood, while the Lummi and Skagit used it to treat tuberculosis, and the Skagit also use it for cuts and sores. Skagit suffering from rheumatism are bathed in water in which very young foliage shoots have been boiled. Both the Lummi and the Skagit chew the pitch-like gum (Gunther 1945).
The western white pine was formerly much desired by lumbermen, especially in the forests of northern Idaho, where it grew to great sizes. Its straight grain makes it a preferred wood for match sticks (Little 1978); though that sounds like a trivial thing, in 1950 Peattie could write of this tree that "every American handles its wood an average of 2300 times a year!" Since it was by far the primary wood used for matches, this use alone consumed 300,000 mature trees per year. Additionally, the white, nonresinous wood is easily worked and cut, both with and across the grain, and takes nails easily without splitting. It was much desired for window and door frames, moldings and trim, pattern stock (heavily used by foundries), cabinetmaking, veneer base, and woodcarving (Arno and Hammerly 2007). All in all, few trees have constituted such a useful and desirable source of lumber. The era of white pine exploitation began early, with the first sawmill built by Rev. Henry Spaulding at his mission in the Nez Perce community of Lapwai (now part of Idaho) in 1840. The market only began to really flourish when the transcontinental railroads reached the northern Rockies, beginning with the Northern Pacific in 1881. This was prior to the establishment of federal forest reserves, besides which Congress had made enormous land grants to the railroads, and intensive logging and clearing of those lands began immediately. The harvest of western white pine reached a rate of 500 million board feet per year from sawmills in the Coeur d'Alene watershed by the early 1900s (Peattie 1950, Arno and Hammerly 2007). At about that time, the Roosevelt administration began setting aside large tracts of land in forest reserves, leading at least to a more regulated exploitation of the species, and a slowing of harvest rates. In 1910, though, two disasters occurred that spelled the end of western white pine as a dominant species in the Rocky Mountains. One was one of the largest forest fires in United States history, centered in northern Idaho's white pine country, and devastating over 3 million acres so completely that the regeneration was dominated mainly by lodgepole pine. This proved to be a pivotal event, triggering a nationwide campaign of fire suppression that was to completely transform the structure and ecology of western U.S. forests for the rest of the 20th century, a story eloquently told by Pyne (2001). Also in 1910, the white pine blister rust was introduced to North America, entering from Asia via the port of Vancouver, BC; that tale is told HERE; suffice it to say that this disease was mortal to the principal white pines of North America, none more so than P. monticola. It took until the 1920's for the blister rust to reach the white pine country of the northern Rockies, but once there, the mortality was almost complete, and white pine virtually disappeared from the forest. A few trees, however, had some resistance to the disease, and survived. Those survivors have been artificially propagated in seed orchards and now millions of resistant white pines have been planted out, and this species is once more becoming a noteworthy component of the forests within its native range (Arno and Hammerly 2007). The tree also has high amenity value, and thus has been widely planted as an ornamental; and at times, collection of the large and attractive cones has been popular (Graham 1990).
Observations
It is the only big pine in western Washington and northwest Oregon, and seems to be easily found in inland coniferous forests throughout that area. It is also reasonably common in the high country of the Sierra Nevada, growing along many popular trails in the national parks and along the Pacific Crest Trail. I have heard that the species' eastern range limit, near the Avalanche Campground in Glacier National Park, is particularly interesting because of the strong resemblance those trees bear to their Pinus strobus counterparts in Minnesota, 1500 km to the east.
Remarks
White pine blister rust (Cronartium ribicola), an introduced fungal disease, has decimated formerly extensive stands of this and certain other white pines (Little 1980).
This species is the principal host for the dwarf mistletoe Arceuthobium monticola (Hawksworth and Wiens 1996).
Western white pine is the state tree of Idaho (Kral 1993).
This is another of the many North American conifers first described by David Douglas, who collected it during his first trip to western North America in 1824-1827, in "the mountains near the base of Mount St. Helens." In his 1827 description, he noted that "This species is intermediate between P. Lambertiana and P. Strobus" and that it "yields a hard, candied, bright amber-coloured rosin," which is consistent with reports that native peoples would chew the gum. He also clearly discerned the two habitats where it is most common, calling it "A handsome tree of large dimensions, particularly in aqueous deposits consisting of decayed vegetable substances in the mountain valleys which are washed by the torrents from higher altitudes, also in rocky, bare, thin soils it particularly abounds" (Douglas 1914, p. 344). See the Topics page for more on Douglas.
Citations
American Forests. 2019. American Forests National Champion Tree List 2018. https://www.americanforests.org/wp-content/uploads/2018/09/American-Forests-Champion-Tree-Register-Updated-Jan-2019.pdf, accessed 2019.07.14.
Arno, Steve and Ramona Hammerly. 2007. Northwest Trees, revised edition. Seattle: The Mountaineers.
Douglas, David. 1914. Journal Kept by David Douglas During His Travels in North America 1823-1827. London: William Wesley & Son. 369pp.
Graham, R. T. 1990. Pinus monticola Dougl. ex D. Don, in Burns and Honkala (1990).
Gunther, Erna. 1945. Ethnobotany of Western Washington. University of Washington Publications in Anthropology 10(1):1-62.
Harlow, W. M., and E. S. Harrar. 1969. Textbook of Dendrology. 5th Edition. McGraw-Hill. pp 313.
Jin, W.-T., D. S. Gernandt, C. Wehenkel, X.-M. Xia, X.-X. Wei, and X.-Q. Wang. 2021. Phylogenomic and ecological analyses reveal the spatiotemporal evolution of global pines. Proc. Natl. Acad. Sci. USA 118(20): e2022302118. doi:10.1073/pnas.2022302118.
Liston, A., M. Parker-Defeniks, J.V. Syring, A. Willyard, and R. Cronin. 2007. Interspecific phylogenetic analysis enhances intraspecific phylogeographical inference: a case study in Pinus lambertiana. Molecular Ecology 16(18):3926-3937.
Price, R. A., A. Liston and S. H. Strauss. 1998. Phylogeny and systematics of Pinus. P.49-68 in Richardson, D.M. (ed.), Ecology and Biogeography of Pinus. Cambridge University Press. ISBN 0-521-55176-5.
Pyne, Stephen J. 2001. Year of the Fires. Viking, 312pp.
Richardson, B.A., G.E. Rehfeldt, and M.-S. Kim. 2009. Congruent climate-related genecological responses from molecular markers and quantitative traits for western white pine (Pinus monticola). International Journal of Plant Sciences 170(9):1120-1131.
Syring, J.V., K. Farrell, R. Businsky, R. Cronin, and A. Liston. 2007. Widespread genealogical nonmonophyly in species of Pinus subgenus Strobus. Systematic Biology 56(2):1-19.
See also
Arno, Stephen F. and Jane Gyer. 1973. Discovering Sierra Trees. Yosemite Natural History Association. 89pp.
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library. This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
FEIS database.
Lanner 1983.
Neuenschwander, Leon F.; Byler, James W.; Harvey, Alan E.; McDonald, Geral I.; Ortiz, Denise S.; Osborne, Harold L.; Snyder, Gerry C.; Zack, Arthur. 1999. White pine in the American West: a vanishing species-can we save it? Gen. Tech. Rep. RMRS-GTR-35. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 20 p.