Podocarpus
L'Heritier ex Persoon 1807
Common names
Yellowwood, brown or black pine.
Taxonomic notes
Syn.: Margbensonia A.V.Bobrov & Melikyan 1998 (POWO, accessed 2023.02.09). Type species, Podocarpus elongatus. Podocarpus is the largest of all conifer genera, here treated in 111 species:
The genus has previously been visited in some depth by Buchholz and Gray (1948a, 1948b, 1948c; Gray 1956, Gray 1958) and by de Laubenfels (1969, 1985), and these authors did excellent work with access to only morphological data, but it was clear that formal cladistic analysis using morphological and molecular markers would be needed to resolve the structure of this large genus. That was finally produced by Knopf et al. (2012) in a landmark study that encompassed 80 Podocarpus taxa (and another 65 taxa of other Podocarpaceae genera) and examined 3 molecular markers, and a comprehensive array of foliar morphology traits. The latter point is particularly important because, as was well shown by Biffin et al. (2011), the evolution of the Podocarpaceae can basically be seen as an evolution of leaves, which in so much of the family look very unlike other conifers and largely account for the family's capacity to persist in warm, wet tropical forests dominated by angiosperm trees. These traits are especially relevant for Podocarpus. Since these studies, important further work has been done on systematics, including recognition of at least 9 new species and acceptance of several others, previously described but unresolved or treated as synonyms; the most extensive of these studies are by de Laubenfels (2015) and Mill (2014, 2015a, 2015b). Nonetheless, the taxonomy of Podocarpus is still less well known than that of any other large conifer genus, and it remains littered with species of doubtful validity, largely due to a long history of erecting new species on the basis of minor morphological variation that may merely reflect phenotypic plasticity or ecotypic variation. It is likely that only more detailed molecular taxonomy will resolve the issue, as it did, for example, in Juniperus and Pinus.
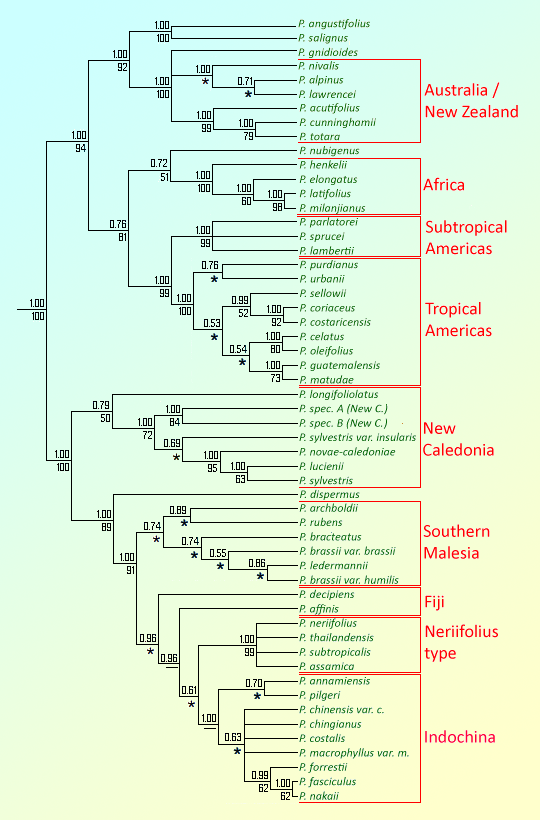
The consensus trees generated by Knopf et al. (2012) vary widely in number of taxa addressed, because not all markers and morphological characters were scored for all species. The consensus trees all agree that species groups are organized primarily by geography, with separate clades for Africa, Australia/New Zealand, New Caledonia, Fiji, southern Malesia, Indochina, subtropical America, and tropical America. The exceptions to this pattern are three "taxonomic" subclades, one allied to the African clade containing P. nubigenus (Chile and Argentina), one allied to the Australian clade containing P. angustifolius (Cuba) and P. salignus (Chile), and the third allied to the southeast Asian subclades, containing a separate group of widely-distributed SE Asian taxa: P. neriifolius, P. subtropicalis, and P. thailandensis (the last treated here under P. neglectus). The analysis supports de Laubenfels' (1985) designation of two subgenera, Podocarpus and Foliolatus, with the former primarily containing African, Australian and American species and the latter primarily containing species of New Caledonia, Fiji, Malesia and Indochina. I have redrafted, from Knopf et al. (2012), the cladogram for Podocarpus which shows combined results for all molecular and morphological markers (shown at right), for 57 taxa.
Description
Dioecious (rarely monoecious), evergreen shrubs and trees to 50 m tall and 350 cm dbh. Leaves alternate, linear to ovate, usually with a single midvein and rarely with parallel veins; leaves bear resin canals, but wood does not. Primary branching in pseudo-whorls, following Massart's model. Terminal foliage buds have imbricate or spreading primary and secondary scales; details of the number, size, and morphology of these buds and scales are often critical to accurate species identification. Leaves spirally inserted or sub-opposite, sessile or petiolate, relatively broad and flattened compared to almost all other conifers, usually linear to lanceolate, variously coriaceous (with more coriaceous leaves typically more long-lived, varying from 1 to >7 years), with a single midrib that is raised or sunken in most species, and flat to strongly revolute margins; the midrib and leaf margins are also important characters in identification. Leaf size is also important, but there is often a gradient in leaf size, with the largest leaves found in shady understory saplings and the smallest leaves in the exposed crown of mature trees; this variability is extreme in some species, and minor in others. Similarly, understory leaves are more likely to bear a drip tip, and sun foliage is more likely to have an obtuse or rounded apex. Stomata are almost always limited to many fine lines on either side of the lower midrib, but in rare instances a few may occur on the adaxial leaf surface. Pollen cones axillary, cylindrical, typically borne in groups in zones along the twig, or peripheral to the terminal bud of the twig; often they have basal bracts called "foliola"; length, diameter, size and shape of foliola, and presence/absence of a peduncle can be useful characters in identification, but many plants are sterile. Pollen cones lengths given are for ripe, active cones; lengths typically increase by 20-30% after pollen has been shed. Microsporophylls triangular, solitary and sessile, or clustered on short sessile or long-pedunculate stalks. Seed cones axillary; receptacle naked or composed of 2-5 bracts, of which the upper 1-2 bracts are sterile and in most species swell greatly to form a succulent, often brightly-colored (orange, red or purple) receptacle; this receptacle functions primarily to attract birds, which are the most important dispersal agent for Podocarpus seeds. Seeds 1(-2), exposed, covered with an often colorful integument called the epimatium; usually references to the "seed" can be taken to mean "the seed with its covering epimatium" (Farjon (2010) and pers. obs.). See also Mill (2015a, p. 70).
Successful identification of species of Podocarpus relies heavily on details of leaf anatomy and foliage bud anatomy, particularly since many plants are sterile or, at least, lack reproductive structures for much of the year. Keys to the species are provided by de Laubenfels (1985) and Farjon (2010), which however do not cover all taxa; adding de Laubenfels (2015) and Mill (2015a, 2015b) largely addresses this issue.
Distribution and Ecology
As discussed above, systematics and geography are closely related in Podocarpus. Like Lepidothamnus, Pectinopitys, Prumnopitys and Retrophyllum, the genus has representatives on both east and west sides of the Pacific Ocean. It thus predates the breakup of Gondwana, which happened in late Cretaceous time when South America separated from Antarctica. The most widespread species is Podocarpus neriifolius; the least widespread are probably P. perrieri, not seen since 1951, and P. victorinianus, not seen since 1924; there is a strong possibility that both are extinct. P. laminaris and P. palawanensis are both known from a single collection and are probably assignable to a different described species, not yet identified.
The following table groups species into the principal geographic regions occupied by the genus. If you need more detail, this document lists all of the countries where each species occurs natively.
| Geography |
Species |
| Africa/Madagascar |
P. capuronii,
P. elongatus,
P. henkelii,
P. humbertii,
P. latifolius,
P. madagascariensis,
P. perrieri,
P. rostratus.
|
| Australia & New Zealand |
P. acutifolius,
P. laetus,
P. dispermus,
P. drouynianus,
P. elatus,
P. grayae,
P. lawrencei,
P. nivalis,
P. smithii,
P. spinulosus,
P. totara.
|
| Central & South America |
P. acuminatus,
P. aracensis,
P. brasiliensis,
P. buchholzii,
P. celatus,
P. costaricensis,
P. glomeratus,
P. guatemalensis,
P. lambertii,
P. magnifolius,
P. matudae,
P. nubigenus,
P. oleifolius,
P. parlatorei,
P. pendulifolius,
P. roraimae,
P. rusbyi,
P. salicifolius,
P. salignus,
P. sellowii,
P. sprucei,
P. steyermarkii,
P. tepuiensis,
P. transiens.
|
| The Caribbean |
P. angustifolius,
P. aristulatus,
P. buchii,
P. coriaceus,
P. ekmanii,
P. hispaniolensis,
P. purdieanus,
P. trinitensis,
P. urbanii,
P. victorinianus.
|
| East Asia |
P. brevifolius,
P. chingianus,
P. fasciculus,
P. hookeri,
P. lenticularis,
P. macrophyllus,
P. nakaii,
P. neglectus,
P. pilgeri,
P. subtropicalis.
|
| Indonesia & Malaysia |
P. archboldii,
P. atjehensis,
P. borneensis,
P. bracteatus,
P. brassii,
P. confertus,
P. crassigemma,
P. deflexus,
P. gibbsiae,
P. glaucus,
P. globulus,
P. laminaris,
P. laubenfelsii,
P. ledermannii,
P. levis,
P. marginalis,
P. micropedunculatus,
P. neolinearis,
P. neriifolius,
P. novoguineensis,
P. oblongus,
P. polystachyus,
P. pseudobracteatus,
P. ramosii,
P. ridleyi,
P. rubens,
P. spathoides,
P. teysmannii,
P. thevetiifolius.
|
| New Caledonia |
P. decumbens,
P. gnidioides,
P. longifoliolatus,
P. lucienii,
P. novae-caledoniae,
P. polyspermus,
P. sylvestris.
|
| Pacific Islands |
P. affinis,
P. costalis,
P. decipiens,
P. insularis,
P. lophatus,
P. macrocarpus,
P. orarius,
P. palawanensis,
P. pallidus,
P. rumphii,
P. salomoniensis,
P. vanuatuensis.
|
Podocarpus occupies very diverse habitats. The most cold-tolerant species occur in temperate climates where snow is unusual and temperatures lower than -10°C are rare. Many taxa are associated with literal islands; the center of diversity for the genus is roughly centered on New Guinea, with high diversity from southern China to New Zealand, and the Caribbean is also a diversity hotspot. Most other taxa are associated with virtual islands in that they are species of the mountains, with a fair number of species endemic to isolated high peaks. A few species, however, occur in lowland tropical rainforest; Podocarpus and Agathis are generally the most successful conifers in this habitat, though a few other conifer taxa are also found there (de Laubenfels 1985, Eckenwalder 2009, Farjon 2010). Podocarpus is often associated with poor soils. On silicate-derived substrates, it is often found in areas where the soils are thin; it is also often competitive on carbonate substrates, on ultramafics, and on excessively leached tropical soils. The extent of adaptation to ultramafic soils is exceptional among conifers, exceeded only by a variety of taxa endemic to New Caledonia (observation based on having documented all the taxa in the genus).
Ecophysiology and the ability of Podocarpus spp. to compete effectively with angiosperms in tropical forest environments have not been widely studied; see P. trinitensis and citations therein for a recent (2021) review of research on this topic. Two recent physiological studies involving P. guatemalensis offer insights to the ecological role of Podocarpus and how that role might change under climate change scenarios. Palma et al. (2020) found that P. guatemalensis and the angiosperm Ficus insipida show comparable growth rates in poor soils, but that a modest addition of fertilizer caused productivity in Podocarpus to double - while increasing it in Ficus by a factor of 20. Most of the difference, they found, could be explained by differences in the hydraulic architecture of Podocarpus and Ficus, such that Ficus could better meet the water demands of rapidly growing tissues. Second, Dalling et al. (2016) found that doubling carbon dioxide concentrations caused a 66% productivity increase in P. guatemalensis while not causing any growth increase in tropical angiosperm trees (they also found a 19% increase in Araucaria heterophylla). These results suggest that the Mesozoic and Paleogene dominance of conifers may have been at least partly due to high atmospheric CO2 concentrations, and also indicate a potential for ongoing anthropogenic CO2 releases to result in a competitive advantage for podocarps in tropical forests.
The podocarps generally do not form extensive stands, instead occurring as individual forest trees (Stevenson 1991). Most examples of Podocarpus as a canopy dominant tree seem to be associated with cloud forests.
Podocarpus is dispersed primarily by birds, which are attracted by the generally bright-colored seeds and their often nutritious receptacles. In reference to South African species: "Seed is dispersed largely by birds. Among these are the brilliantly plumed Loerie of the forests and the Rameron Pigeon that scatter the seed of many kinds of trees and so play a vital part in the life of the forests. Yellowwoods largely control the distribution of a particularly interesting species of bird, the Cape parrot, Poisephalus robustus, in the eastern districts. These birds normally roost and nest in the highest mountain forests of the south east Cape, from where they visit the surrounding country in search of food, principally the kernels of the nuts of the yellowwood fruits" (Palmer and Pitman 1972).
Remarkable Specimens
The biggest tree in the genus is the totara, Podocarpus totara. The tallest is very uncertain, with specimens in both Malesia and South America thought to exceed 50 m tall.
The oldest known Podocarpus is documented by a ring count on an unspecified South African species (probably P. latifolius but maybe Afrocarpus falcatus), 700 years (Palmer and Pitman 1972). There is also a radiocarbon date on a tree core from a P. totara of 890 years, although radiocarbon dates from trees with rotten wood are always suspect due to the possibility of translocation of carbon within the organism, e.g. by fungi. Another ring count on P. laetus was 496 years, and another ring count on P. totara was 460 years. The paucity of data is likely related to the difficulty of aging many of the tropical species, as discussed below (Ethnobotany). For all other species, no data. Given that some of the species are large trees in late-successional forests, I would not be surprised if somewhere out there is a tree more than 1,000 years old.
Ethnobotany
Human use is largely confined to relatively large individuals that grow in proximity to populated areas. Since many of the species in the genus include small trees with very limited distribution, often at high elevation, the majority of taxa have no recorded human use.
Aboriginal and historical uses: Ethnopharmacological uses of bark and leaf extracts are reported for many species, to treat a variety of ailments. The wood of nearly all species is strong, straight-grained, relatively easy to work and used for purposes such as construction, boat-building, furniture and household implements.
Economic uses: In reference to South African species (P. elongatus, P. henkelii, P. latifolius and Afrocarpus falcatus): "Yellowwoods played a very important role in the early life of the colony for their timber was the most generally useful of any found in the country. Although the wood does not weather well and has never, therefore, been much used for exterior doors and windows, it was once popular for indoor work, for ceilings and floors, and for furniture. Its fine yellow colour is now much admired. It did not, however, suit 19th century taste and during this period it was frequently painted. Today the wood is often used to make butchers' blocks because it is hard, without scent, and does not chip easily. Coffins were once often made of it, and sometimes still are. On account of their usefulness, yellowwoods have been some of our most heavily exploited trees" (Palmer and Pitman 1972). The same can be said for many other species that achieve large size; where such trees occur, such as in Australia and parts of east Asia, Malesia, and South America, Podocarpus remains a popular and economically significant timber. However, in much of its range, large trees have been depleted by timber harvest and habitat loss, and due to slow growth rates the species has lost its former economic significance.
Research uses: There has been limited use of Podocarpus in dendrochronological research. Studying four species in Thailand, Buckley et al. (1995) reported that "While most of the [Podocarpus species] appeared to have many hundreds of rings, the ring boundaries were often indistinct, and poor circuit uniformity and wedging of rings was observed." Similar problems have also been reported in P. lawrencei, a temperate alpine species of Australia. If these circumstances are common in the genus, then dendrochronological potential is poor. However, some successful work has been done with the temperate species of New Zealand, and other temperate species likely warrant investigation. Many Podocarpus species have been investigated for pharmacological uses and leaf and bark extracts have been shown to have activities antimicrobial, antiinflammatory, and in other ways useful. Thus far, though, no species has proven essential for production of a medicine.
Observations
See the species accounts.
Remarks
"Podocarpus is derived from two Greek words pous= foot, and karpos= fruit, referring to the fleshy fruit stems" (ANBG [no date]).
On 1492.11.25, Christopher Columbus wrote that the pines he found in the mountains of Cibao, Hispaniola, were not true pines because they did not bear ‘pine cones’ but instead had fruits resembling olives. Consequently, Columbus is credited by Alexander von Humboldt as having distinguished what are now Podocarpus and Pinus long before L’Héritier in 1807 (Mill 2015a).
Citations
[ANBG] Australian National Botanic Gardens. [no date]. Australian Conifers. http://www.anbg.gov.au/anbg/conifers/, accessed 2002.11.09, now defunct.
Buckley, Brendan M., Mike Barbetti, Manas Watanasak, Rosanne D'Arrigo, Saran Boonchirdchoo, and Sakunyut Sarutanon. 1995. Dendrochronological investigations in Thailand. IAWA Journal 16(4):393-409.
Laubenfels, David J. de. 2015. New sections and species of Podocarpus based on the taxonomic status of P. neriifolius (Podocarpaceae) in tropical Asia. Novon 24(2):133-152. https://doi.org/10.3417/2012091.
Dalling, James W., Lucas A. Cernusak, Klaus Winter, Jorge Aranda, Milton Garcia, Aurelio Virgo, Alexander W. Cheesman, Andres Baresch, Carlos Jaramillo, and Benjamin L. Turner. 2016. Two tropical conifers show strong growth and water-use efficiency responses to altered CO2 concentration. Annals of Botany 118(6):1113-1125.
Mill, R. R. 2014. A monographic revision of the genus Podocarpus (Podocarpaceae): I. Historical review. Edinburgh Journal of Botany 71 (3): 309–60. https://doi.org/10.1017/S0960428614000146.
Mill, R. R. 2015a. A monographic revision of the genus Podocarpus (Podocarpaceae): II. The species of the Caribbean bioregion. Edinburgh Journal of Botany 72 (1): 61–185. https://doi.org/10.1017/S0960428614000328.
Mill, R. R. 2015b. A monographic revision of the genus Podocarpus (Podocarpaceae): III. The species of the Central America and northern Mexico bioregions. Edinburgh Journal of Botany 72 (2): 243–341. https://doi.org/10.1017/S0960428615000050.
Palma, Ana C., Klaus Winter, Jorge Aranda, James W. Dalling, Alexander W. Cheesman, Benjamin L. Turner, and Lucas A. Cernusak. 2020. Why are tropical conifers disadvantaged in fertile soils? Comparison of Podocarpus guatemalensis with an angiosperm pioneer, Ficus insipida. Tree Physiology 40(6):810-821.
See also
Enright, Neal J. and Robert S. Hill. 1990. Ecology of the southern conifers. Washington, DC: Smithsonian Institute Press. 342pp.
Turner, B. J. and L. A. Cernusak (eds.). 2011. Ecology of the Podocarpaceae in tropical forests. Washington, DC: Smithsonian Institution Scholarly Press. vii+207.