Pinus nigra
J.F. Arnold 1785
Common names
European black pine, Austrian pine, Cevennes black pine, Corsican pine, Crimean pine, Spanish black pine [English]; Pin laricio [French]; Pinu lariciu [Corse], Schwarzkiefer, Schwarzföhre [German]; Cheren Bor [Bulgarian]; Crni Bor [Serbian]; Karaçam [Turkish]. As discussed below, the species consists of a number of geographically distinct taxa, so many of these common names are typically applied to only one of these taxa.
Taxonomic notes
This is a member of Pinus sect. Pinus subsect. Pinus, within which its position remains unresolved (Gernandt et al. 2005). I here follow the classification of Farjon (2010), under which there are five geographically distinct subspecies and one nothospecies:
- Pinus nigra subsp. nigra;
- Pinus nigra subsp. dalmatica (Vis.) Franco 1943;
- Pinus nigra subsp. laricio (Poir.) Maire 1928;
- Pinus nigra subsp. pallasiana (Lamb.) Holmboe 1914;
- Pinus nigra subsp. salzmannii (Dunal) Franco 1943; and
- Pinus × neilreichiana Reichardt 1876
Like the other two widespread European pines, P. sylvestris and P. mugo, this species has received a vast number of described names, a consequence of very narrow subspecific and varietal concepts applied notably in France and parts of Eastern Europe (I here list only the more prominent synonyms). Although it is variable, it is not markedly so compared to other widely distributed conifers worldwide; across its entire range, it shows a level of genetic diversity similar to many other Pinus species (Scaltsoyiannes et al. 1994). The most prominent variation in P. nigra occurs between trees from the eastern and western halves of its range, which differ strongly in foliage (Christ 1863, Koehne 1893, Delevoy 1949, Dallimore et al. 1967, Vidakovic 1974, Christensen 1993) and genetic structure (Scaltsoyiannes et al. 1994). The two halves of the species' range are separated by a large range gap across the Adriatic Sea and the Po valley of Northern Italy, leaving unoccupied over 600 km of apparently suitable habitat in the southern Alps. These two large areas account for the primary subspecies, subsp. nigra in the east and subsp. salzmannii in the west. Additional distinct subspecies are primarily associated with islands: subsp. dalmatica on the coast of Croatia and adjacent islands; subsp. laricio on Corsica; and subsp. pallasiana on Cyprus and along part of the Black Sea coast. Pinus × neilreichiana occurs in Austria.
Pinus nigra subsp. nigra: Primary synonym P. nigra subsp. croatica Lovric 1972. The type is the illustration in Arnold (1785).
Pinus nigra subsp. dalmatica (Vis.) Franco 1943: Primary synonyms are P. dalmatica Vis. 1842 and P. nigra var. dalmatica Buskinsky 2008. Type not designated.
Pinus nigra subsp. laricio (Poiret) Maire 1928: Primary synonyms include P. laricio Poiret ex Lamarck 1804, non Savi 1798; P. nigra var. corsicana (Loudon) Hylander; P. nigra var. maritima (Aiton) Melville; var. P. nigra calabrica (Loudon) Schneider; P. nigra var. laricio (Poiret) Maire; P. nigra var. poiretiana (Antoine) Schneider. The varietal name maritima, though widely used and older than corsicana, is invalid. Its type, an illustration cited by Aiton under his Pinus sylvestris var. maritima, proved to be Pinus halepensis, so this name cannot be used for the Corsican pine (Christensen 1993). Type not designated.
Pinus nigra subsp. pallasiana (Lamb.) Holmboe 1914: Primary synonym is P. pallasiana Lamb. 1824, also P. pontica K. Koch. The type is the illustration in Lambert (1824); presumably the type specimen cited by the author as in Cyprus, "Above Prodromo (JH 955)" (Holmboe 1914) is lost.
Pinus nigra subsp. salzmannii (Dunal) Franco 1943: Primary synonyms are P. salzmannii Dunal 1851; P. nigra var. salzmannii (Dunal) Laguna Lumbreras 2000; P. nigra var. cebennensis (Godron) Rehder; P. nigra var. monspeliensis (Salzmann) Slavin; P. nigra var. tenuifolia (Parl.) Schneider; P. pyrenaica Lapeyrouse; P. clusiana Clemente; P. nigra var. mauretanica Maire et Peyerimhoff; and P. nigra ssp. mauretanica (Maire & Peyerimhoff) Heywood. Type: France, Languedoc, Hérault, St. Guillaume-le-désert, P. salzmann s.n., holotype MPU.
Pinus × neilreichiana Reichardt 1876: The natural hybrid between P. nigra subsp. nigra and Pinus sylvestris. No synonyms. Type: Austria, Niederösterreich, Baden, Grossau, along footpath to Pottenstein, Reichardt s.n., lectotype C.
Description
Trees to 50 m tall and 190 cm dbh, usually with a single round trunk and ascending branches forming a broad-conical to domed crown that flattens with age. Bark on mature trees thick, breaking into scaly ridges or irregular plates, deeply fissured, dark gray, can be pinkish on some old trees. Twigs stout, rough with pulvini from fallen fascicles, new shoots yellow-green, soon darkening during first summer to red-brown. Buds ovoid-acuminate, red-brownish with whitish fringes to scales, usually with some patchy grey-white resin. Leaves in fascicles of 2 with a persistent 10-12 mm long sheath, straight to curved, stiff to flexible, (4-)8-16(-18) cm long and 1-2 mm thick, light to dark green, edges minutely serrate, apex acute, stomata in lines on both faces. Pollen cones clustered at base of current year's shoots, ovoid-conical to cylindrical, 15-25 × 5-7 mm, yellow when ripe. Seed cones solitary or in whorls of 2-5, borne on short stalks, when closed conical-ovoid, (3.5-)5-10(-12) × 2-4 cm, ovoid when open, light brown, falling soon after seeds are released. Cone scales thin, woody, rigid; apophyses slightly raised and transversely keeled; umbo dorsal, small, unarmed or with a tiny deciduous prickle. Seeds flattened, obovoid, 6-8 mm long, gray, often mottled, with a 15-25 mm light brown wing (Frankis emails 1999, Farjon 2010). Pollination April-May, cones mature 20 months later in October-December, and open to shed seed from February-April (Frankis email 1999).
The subspecies may be distinguished as follows (Farjon 2010):
- P. nigra subsp. nigra: Trees to 40 m tall. Bark dark gray, breaking into longitudinal ridges and fissures and small scaly plates. Leaves stiff, curved, 8-13 cm long, dark green.
- P. nigra subsp. dalmatica: Trees to 15 m tall. Leaves 4-7 cm long. Seed cones (3-)4-6(-7) cm long.
- P. nigra subsp. laricio: Trees to 50 m tall. Bark gray with deep longitudinal fissures and irregular scaly plates. Leaves slender, flexible, 8-15 cm long and 1.2-1.5 mm thick, light green.
- P. nigra subsp. pallasiana: Trees to 40 m tall, often with paired or forked trunks. Bark light gray with deep longitudinal fissures and relatively large scaly plates. Leaves stiff, straight or curved, (8-)12-17 cm long, light green. Some trees, that have been described as P. nigra subsp. pallasiana var. fastigiata Businsky 1999, have a fastigiate growth form.
- P. nigra subsp. salzmannii: Trees to 30 m tall. Bark light brown, variously fissured, with irregular, scaly plates. Leaves flexible, 8-16 cm long and 1-1.3 mm wide.
P. × neilreichiana most closely resembles P. nigra subsp. nigra, but it has reddish bark on the branches and nearly flat apophyses on the cone scales. It differs from P. sylvestris in having longer, thicker, dark green leaves (Farjon 2010).
Besides the differences noted above, there are also some consistent differences in details of leaf anatomy; see Farjon (2010) for details.
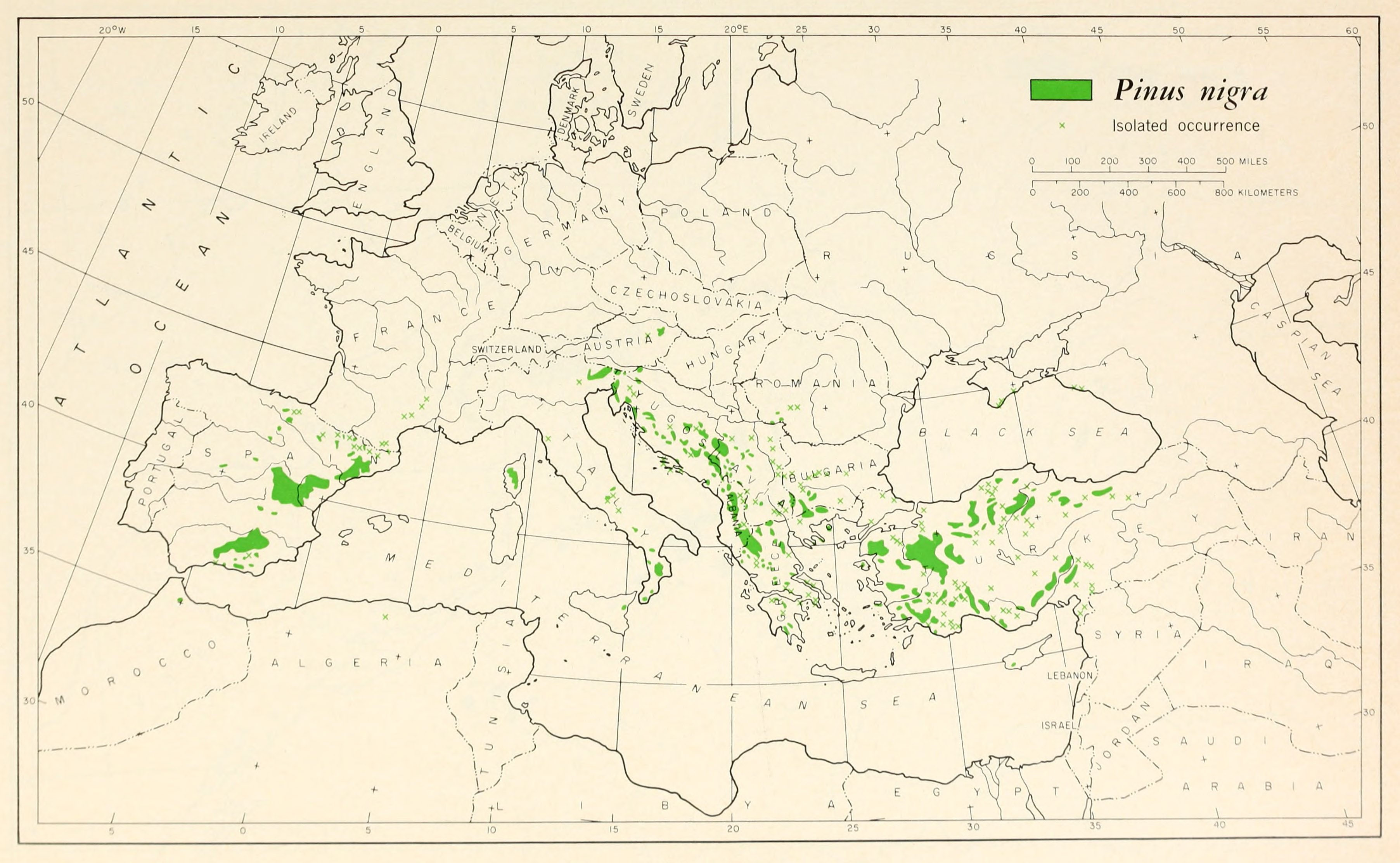
Distribution and Ecology
Native to Albania, Algeria, Andorra, Austria, Bosnia and Herzegovina, Bulgaria, Croatia, Cyprus, France, Greece, Italy, Macedonia, Montenegro, Morocco, Romania, Russia, Slovenia, Spain, Turkey, and Ukraine (Conifers of the World). Naturalized in certain other countries, e.g. Australia, Canada (Alberta, British Columbia, Ontario, and Québec), Great Britain, New Zealand, Portugal, and the United States (Illinois, Maine, Massachusetts, Michigan, Missouri, New Jersey, New York, Ohio, Pennsylvania, and probably Washington).
This widely distributed species has diverse and complex environmental correlates. The picture is even more complex because it is not always clear if a population is natural or has been established and maintained, at least in part, through human agency; it has long been a mainstay of Europe's timber industry. P. nigra has varied soil tolerances, from acidic to basic. It is generally a lower montane species. It sometimes form pure stands, but is often in mixed stands (especially with P. sylvestris), and locally occurs with a very wide array of other trees, both conifers and hardwoods. It is more tolerant of salt than P. sylvestris and often occurs near the sea; this salt tolerance also varies between taxa, with subsp. laricio substantially more tolerant than subsp. nigra (Farjon 2010).
The difference in foliar morphology between the subspecies reflects a more severe winter climate in the eastern populations. Winter temperature minima are markedly lower in this more continental climate, with absolute minima sometimes falling below -30°C in most of the eastern subspecies' range. Minima in the western subspecies' range rarely fall as low as -20°C. Forestry trials in cold areas of the USA (Lee 1968, Wheeler et al. 1976) have consistently found eastern origins to be significantly more cold-tolerant than western origins (Frankis emails 1999).
Distribution and ecology for the infraspecific taxa are as follows:
P. nigra subsp. nigra
Native to Albania, Austria, Bosnia and Herzegovina, Bulgaria, Croatia, Greece, Macedonia, Montenegro, Romania, Serbia, Slovenia, and Turkey (Farjon 2013, accessed 2019.02.26). Generally found at elevations of 200-1200 m, in southeast European continental climates with some summer rainfall (Frankis emails 1999). Not of conservation concern.
P. nigra subsp. dalmatica
Native to Croatia (Conifers of the World), where it occurs at 400-700 m elevation on three islands, none larger than 300 km², and a peninsula of similar size. Within these, the stands are again limited, with four locations known on the southernmost island. The population is severely fragmented, with continuing decline of mature trees, due largely to habitat degradation by feral goats. There are no conservation measures in place to reduce these threats. For these reasons, it has been classified as Endangered (Farjon 2013, accessed 2019.02.26).
P. nigra subsp. laricio
Native to France (Corsica) and Italy (Sicily, on Mt. Etna; and the Apennines in Calabria), at elevations of 950-1800 m. Within its range, the species is widespread, although locally subject to logging or habitat loss. Several major stands, in Corsica, Calabria, and Sicily, are in protected areas. For these reasons, it has been classified as Least Concern (Farjon 2013, accessed 2019.02.26).
P. nigra subsp. pallasiana
Native to Cyprus, Russia, Turkey, and Ukraine, at elevations of 100-1900 m. This has the greatest extent of occurrence of all the subspecies, although it is severely fragmented, and is classified as Least Concern for conservation purposes. It mainly occurs in Turkey, where large stands remain in the Pontic and Taurus Mountains; in the Taurus it forms pure stands, or is mixed with Cedrus libani. In interior Anatolia, in mainly occupies north-facing mountain slopes and ravines (Farjon 2013, accessed 2019.02.26).
P. nigra subsp. salzmannii
Native to Algeria (Montagnes du Hodna), Andorra (presumed), France (Cevennes and Pyrenees Mountains), Morocco (Rif Mountains), and Spain, at elevations of 400-1500 m. It is widespread in Spain, both as natural stands and in plantation forestry. The North Africa populations are small and isolated, and probably genetically distinct, but as currently circumscribed the taxon is classified as Least Concern for conservation purposes. It typically occurs in low mountains or hills, on sandy or rocky soils with good drainage (Farjon 2013, accessed 2019.02.26).
P. × neilreichiana
Native to Austria (Conifers of the World). No other environmental description found.
P. nigra is stated to be hardy to Zone 5 (cold hardiness limit between -28.8°C and -23.3°C) (Bannister and Neuner 2001), but the subspecies is not stated. Presumably it was not an African population.
Remarkable Specimens
The largest tree in habitat, and second-largest known, appears to be a specimen of subsp. laricio growing in the vallée de la Restonica in Corte, Haute-Corse, France; when measured in 2008 it was 600 cm girth (191 cm dbh) with an estimated height of 22 m (Monumental Trees 2020). The largest diameter specimen, of ssp. pallasiana, grows in the garden of a private house in Dallinghoo, Suffolk, UK; it is reported to be 730 cm girth (232 cm dbh) with an estimated height of 27.0 m when measured in 2018 (Monumental Trees 2018). No photo is provided.
Kouta Räsänen (2012) reports that the Durmitor National Park in Montenegro has many pines 45 m tall, with the highest measured being 47.4 m (laser measurement) and 94 cm dbh (photos at Monumental Trees). He measured other pines at Durmitor up to 133 cm dbh. Farjon (2010) reports trees to 50 m tall and 189 cm dbh in Corsica, but provides no further details. A 45 m tall tree, probably subsp. laricio, was measured in the Atlantic Pyrenees by Dominique Beziat (e-mail 2020.11.29) using a laser, in 2018. A tree 185 cm dbh and 43 m tall is known from Fallistro, Calabria (Corpo Forestale della Stato 2004). Generally the Corsican pines (subsp. laricio are renowned for their large size, and it is reasonable to expect that they include the largest specimens in this species.
A comprehensive review of this species, including many trees of extraordinary size and height, is given at MonumentalTrees.com.
The largest known tree in North America, shown here, is 121.3 cm DBH and 31.2 m tall, with a 16.8 m crown diameter, measured in 2018.03 (Robert Van Pelt email 2018.04.01).
This species has been the subject of innumerable dendrochronological collections, and old trees with ages verified by crossdating and archived wood have been found widely dispersed through its range. The most notable examples include:
- 849 years for a living tree in Corsica, France, collected by P. Römer and colleagues (doi.org/10.25921/09mc-nm03). This chronology was used in a long-term temperature study (Römer et al. 2021).
- 800 years for a dead tree and 668 years for the oldest living tree, in the Sierra de Cazorla of Spain. Chronology collected by Emilia Gutiérrez (doi.org/10.25921/jsa2-6b42) and used in a climate change study (Shestakova et al. 2019). The Sierra de Cazorla has also yielded a living tree with an 803-year record (Tíscar and Lucas-Borja 2016).
- 688 years for a dead tree and 603 years for the oldest living tree, in the Ilgaz Mountains of northern Turkey. Chronology collected by Nesibe Köse and H. Tuncay Güner (doi.org/10.25921/554n-6n93) and used in a study of long-term temperature variation (Köse et al. 2017).
- 624 years for a living tree in Cyprus, collected by R. Touchan and M. K. Hughes (doi.org/10.25921/bajm-rk11) and used in a study of long-term precipitation variation (Touchan et al. 2005).
Ethnobotany
Subspecies laricio, nigra and pallasiana are widely used in Europe and the US as ornamentals, and in forestry for shelterbelts, and for timber production. Subspecies salzmannii is used for timber production in Spain. P. nigra is generally very tolerant of chalk and limestone, and also of urban pollution - perhaps the most pollution-tolerant of any pine (Frankis emails 1999).
The species has been used in dendrochronology, e.g. a study of the climate-growth response of subsp. salzmannii in southeastern Spain (Martín-Benito et al. 2008), a study of possible climate change response in southern Spain (Linares and Tíscar 2010), and a fire history study in southern Greece (Christopoulou et al. 2013). A study of stand age and structure in an old-growth forest in southern Spain is particularly interesting, both for the detailed analysis and because it just sounds like an extraordinary stand (Tíscar and Lucas-Borja 2016).
Observations
Crna Poda in Durmitor National Park, Montenegro, may be the finest remaining old-growth P. nigra (subsp. nigra) forest. Besides P. nigra, it also hosts fine stands of Abies alba, Picea abies; and even Pinus mugo on the high peaks (Räsänen 2012).
The "Poyo de Santo Domingo" forest, located at the southern limit of the Cazorla, Segura y Las Villas Parque Natural in Spain, is the home of the oldest known tree in the species. For details, see Tíscar and Lucas-Borja (2016).
Remarks
The epithet nigra recognizes the dark gray to nearly black bark of the type subspecies; when found growing with P. sylvestris, the contrast is striking. The epithet neilreichiana remembers German botanist August Neilreich (1803-1871) (Farjon 2010).
Citations
Arnold, J. F. 1785. Reise nach Mariazell in Steyermark. Wien (p. 8).
Christ, H. 1863. Uebersicht der Europäischen Abietineen (Pinus Linn.). Verhandlungen der Naturforschenden Gesellschaft Basel (n.s.) 3: 541-557.
Christensen, K. I. 1993. Comments on the earliest validly published varietal name for the Corsican Pine. Taxon 42: 649-653.
Christopoulou, A., P. Z. Fulé, P. Andriopoulos, D. Sarris, and M. Arianoutsou. 2013. Dendrochronology-based fire history of Pinus nigra forests in Mount Taygetos, southern Greece. Forest Ecology and Management 293:132–139.
Critchfield, William B. and Elbert L. Little. 1966. Geographic Distribution of the Pines of the World. USDA Forest Service Miscellaneous Publication 991.
Corpo Forestale della Stato. 2004. Alberi Monumentali D'Italia. http://www.corpoforestale.it/foreste&forestale/ricerca&progetti/alberi_m/index.htm, accessed 2004.12.08.
Delevoy, G. 1949. A propos de la systématique de Pinus nigra Arnold. Travaux Station Recherches de Groenendaal série B. 12: 1-37.
Farjon, Aljos. 2010. A Handbook of the World's Conifers. Leiden, Netherlands: Brill Academic Publishers.
Franco, João do Amaral. 1943. Dendrologia Florestal. Lisboa: Lucas. pp. 55 and 56.
Gernandt, D. S., G. Geada López, S. O. Garcia and A. Liston. 2005. Phylogeny and classification of Pinus. Taxon 54(1):29-42.
Holmboe, Jens. 1914. Studies on the vegetation of Cyprus. Bergens Museums Skrifter ser. 2 1(2). Page 29, accessed 2019.02.26.
Koehne, B. A. E. 1893. Deutsche Dendrologie. Stuttgart.
Köse, Nesibe, H. Tuncay Güner, Grant L. Harley, and Joel Guiot. 2017. Spring temperature variability over Turkey since 1800 CE reconstructed from a broad network of tree-ring data. Climate of the Past 13(1):1-15. doi: 10.5194/cp-13-1-2017.
Lambert, Aylmer Bourke. 1824. A description of the genus Pinus, illustrated with figures, directions relative to the cultivation, and remarks on the uses of the several species. Vol. 2. London: J. White. P. 1, accessed 2019.02.26.
Lee, C. H. 1968. Geographic variation in European Black Pine. Silvae Genetica 17: 165-172.
Linares, J. C., and P. A. Tíscar. 2010. Climate change impacts and vulnerability of the southern populations of Pinus nigra subsp. salzmannii. Tree Physiology 30(7):795–806.
Maire, R. 1928. Contribution à l'étude de la flore de l'Afrique du Nord. Bulletin de la Société d'histoire naturelle de l'Afrique du nord 19. Page 66, accessed 2019.02.26.
Martín-Benito, D., P. Cherubini, M. del Río, and I. Cañellas. 2008. Growth response to climate and drought in Pinus nigra Arn. trees of different crown classes. Trees 22(3):363–373.
Mitchell, A.F., V.E. Hallett, and J.E.J. White. 1990. Champion trees in the British Isles. Forestry Commission Field Book 10.
Räsänen, K. 2012. Crna Poda – a new height record for European black pine. www.ents-bbs.org/viewtopic.php?f=385&t=4725, accessed 2013.02.24. The article includes many photographs.
Reichardt. 1876. Verhandlungen der Zoologisch-Botanischen Gesellschaft in Wien V. 26, P. 461-463, accessed 2019.02.26.
Römer, Philipp, Claudia Hartl, Lea Schneider, Achim Bräuning, Sonja Szymczak, Frédéric Huneau, Sébastien Lebre, Frederick Reinig, Ulf Büntgen, and Jan Esper. 2021. Reduced temperature sensitivity of maximum latewood density formation in high-elevation Corsican pines under recent warming. Atmosphere 12(7), doi: 10.3390/atmos12070804.
Scaltsoyiannes, A., Rohr, R., Panetsos, K. P., & Tsaktsira, M. 1994. Allozyme frequency distributions in five European populations of Black Pine (Pinus nigra Arnold). Silvae Genetica 43: 20-30.
Shestakova, Tatiana A., Jordi Voltas, Matthias Saurer, Frank Berninger, Jan Esper, Laia Andreu-Hayles, Valérie Daux, Gerhard Helle, Markus Leuenberger, Neil J. Loader, Valérie Masson-Delmotte, Antonio Saracino, John S. Waterhouse, Gerhard H. Schleser, Zdzislaw Bednarz, Tatjana Boettger, Isabel Dorado-Liñán, Marc Filot, David Frank, Michael Grabner, Marika Haupt, Emmi Hilasvuori, Högne Jungner, Maarit Kalela-Brundin, Marek Krapiec, Hamid Marah, Slawomira Pawelczyk, Anna Pazdur, Monique Pierre, Octavi Planells, Rutile Pukiene, Christina E. Reynolds-Henne, Katja T. Rinne-Garmston (Rinne), Angelo Rita, Eloni Sonninen, Michel Stiévenard, Vincent R. Switsur, Elzbieta Szychowska-Krapiec, Malgorzata Szymaszek, Luigi Todaro, Kerstin Treydte, Adomas Vitas, Martin Weigl, Rupert Wimmer, and Emilia Gutiérrez. 2019. Spatio-temporal patterns of tree growth as related to carbon isotope fractionation in European forests under changing climate. Global Ecology and Biogeography 28(9):1295-1309. doi: 10.1111/geb.12933.
Tíscar, P. A., and M. E. Lucas-Borja. 2016. Structure of old-growth and managed stands and growth of old trees in a Mediterranean Pinus nigra forest in southern Spain. Forestry: An International Journal of Forest Research 89(2):201–207.
Touchan, R., E. Xoplaki, G. Funkhouser, J. Luterbacher, M. K. Hughes, N. Erkan, Ü. Akkemik, and J. Stephan. 2005. Reconstructions of spring/summer precipitation for the eastern Mediterranean from tree-ring widths and its connection to large-scale atmospheric circulation. Climate Dynamics 25:75-98.
Vidakovic, M. 1974. Genetics of European Black Pine (Pinus nigra Arnold). Annales Forestales 6/3: 57-86.
Wheeler, N. C., Kriebel, H. B., Lee, C. H., Read, R. A., & Wright, J. W. 1976. 15-Year performance of European Black Pine in provenance tests in North Central United States. Silvae Genetica 25: 1-6.
See also
Dida, M., F. Ducci, and G. Zeneli. 2001. Black pine (Pinus nigra Arn.) resources in Albania, in Forest Genetic Resources No. 29. www.fao.org/DOCREP/004/Y2316E/y2316e0d.htm, accessed 2011.02.25.
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library (as P. laricio). This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
Gülsoy, A. D., A. M. Gülsoy, B. çEngel, and Z. Kaya. 2014. The evolutionary divergence of Pinus nigra subsp. pallasiana and its varieties based on noncoding trn regions of chloroplast genome. Turkish Journal of Botany 38:627–636.
Maire, R. and P. de Peyerimhoff. 1927. Sur la découverte d'un Pin Laricio dans l'Afrique du Nord. Comptes Rendus 184: 1514-1516.
Mitchell, A.F. 1972. Conifers in the British Isles. Forestry Commission Booklet 33.
Nikolic, D., and N. Tucic, N. 1983. Isoenzyme variation within and among populations of European Black Pine (Pinus nigra Arnold). Silvae Genetica 32: 80-89.