Conservation Status

Agathis australis
(D. Don) Lindl. ex Loudon 1829
Common names
Kauri, New Zealand kauri.
Taxonomic notes
Syn: Dammara australis (D. Don) Lamb. 1824, Podocarpus zamiifolius A. Rich. 1832 (Farjon 1998).
Description
Monoecious trees 30-50 m tall and 100-400 cm or more in diameter. Young trees typically dominate their stands and have narrowly columnar crowns, which, after emerging above the codominant forest canopy, become globular or flat-topped. Trunk remarkably cylindrical, with almost no visible taper, often 15 m or more to the first branch on large trees. Bark ash-grey, blue-grey, or mottled purple, falling in large, thick flakes. Branches in whorls or irregularly horizontal; branches are shed at a basal abscission layer soon after their foliage dies, emphasizing the cylindrical bole form. The abscised small branchlets are typically a major component of the understory litter. Branchlets are smooth, glaucous. Buds are rounded, with imbricate scales. Leaves are alternate to subopposite, sessile, thick, coriaceous, parallel-veined. Juvenile leaves are glaucous-green, lanceolate, 5-10 cm × 5-12 mm. Adult leaves are green, slightly glaucous beneath near the petiole, ovate-lanceolate, 2-3.5 cm × 10-15 mm, apex obtuse, with 15 parallel nerves and a short petiole. Pollen cones are stout, cylindrical, 2-5 cm long, on a stout peduncle 1 mm wide; sporophylls are imbricate, margin weakly erose. Female cone subglobose, on a short peduncle, 5-7.5 cm diameter, glaucous-green, scales 1.8 cm long with a short mucronate umbo (Allan 1961, Silba 1986, pers. obs. Mar-2003). The reproductive cycle extends over 19-20 months from pollination in October to seed maturity in February or March of the second year following (Owens et al. 1997). See García Esteban et al. (2004) for a detailed characterization of the wood anatomy.
"A tree twenty-five to thirty years old may begin to bear fertile seeds. When the female cone is ripe the scales open, releasing the winged seeds which are borne away by the wind. Seeds remain viable for only a short period, and usually germinate when they settle in an open situation where light can penetrate readily.
"The spreading crown of the kauri is, relative to the trunk, immense; it is rather open as the tree ages and is supported on huge branches that appear simply to swell out of the upper trunk. The first or lower of these branches can be up to 2 m through where they leave the trunk. Kauri trees are rather slow growing over their life span although young trees have been recorded as growing at the rate of 0.35 m per year. ... Young kauri trees ... are known as "rickers". They persist in this juvenile form for fifty years or more and can attain a considerable height with their tall thin trunks and narrow pyramid shaped crowns" (Salmon 1996).
Distribution and Ecology
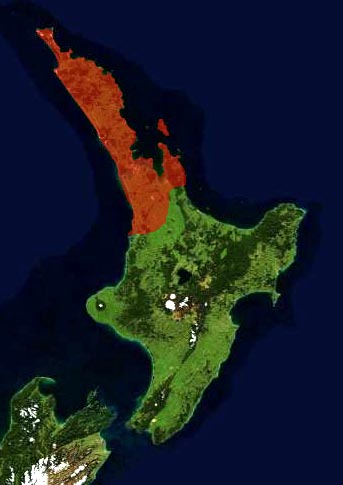
New Zealand: N Island, Northland and Coromandel Peninsula, south to about lat. 38°, the southern limit was bounded by a line from Raglan Harbour through Hamilton to a little south of Tauranga (Salmon 1996). It grows primarily in mountains at 150-400 m elev. but is generally found at 0-600 m elevation. Lives in a forest that commonly includes totara (Podocarpus totara), montane totara (Podocarpus laetus), rimu (Dacrydium cupressinum), and miro (Pectinopitys ferruginea).
The conservation status of this species has not been assessed by the IUCN, but the New Zealand Threat Classification System assesses it as "Nationally Vulnerable" (accessed 2021.11.25).
"In even the oldest trees the trunk is kept clean and free of epiphytes by the continuous shedding of small and large flakes of bark which build up in a large mound, 2 m or more in height, around the base of the tree. This, in time, decays to rich humus and is densely penetrated by the roots of the tree, giving the impression that the tree is growing from it. Kauri bark is fairly thick and exudes gum, which also flows copiously from any wound to the tree as well as from cones and even, at times, from branchlets and the bases of leaves" (Salmon 1996).
"No fossil evidence has been found of any earlier existence of kauri forests south of their present-day occurrence" Salmon 1996.
An account of the kauri dominated ecosystems at Waipoua Forest has been published by Burns and Leathwick (1996). They note that the forest occurs within the 13,000 ha Waipoua Forest Sanctuary and Waipoua Kauri Management and Research Area. This is the largest relatively pristine forest left in the Northland; it extends 19 km inland from the coast and rises to an elevation of 640 m, some of the highest country in the Northland. Deeply weathered, infertile clay soils overlie Waipoua basalt bedrock; annual rainfall is about 1650 mm/year at sea level, rising to about 2500 mm/year at the highest elevations. The kauris mainly occur in discrete stands, usually located on relatively infertile ridgetop soils, although rimu (Dacrydium cupressinum) is an emergent dominant in the highlands. Intervening forests are dominated by taraire (Beilschmiedia tarairi) and kohekohe (Dysoxylum spectabile). Heathlike scrubs are locally extensive at lower altitudes. Kauri regeneration chiefly occurs in gaps. Burns and Leathwick hypothesize that the distribution of kauri on the landscape is partially the result of a feedback whereby the acidic decomposition of conifer litter leaches the poor soils further, perpetuating conifer dominance in areas already having conifer stands.
Hardy to Zone 9 (cold hardiness limit between -6.6°C and -1.1°C) (Bannister and Neuner 2001).
Kauri dieback is a growing problem in New Zealand. This is an introduced disease caused by a Phytophthora (in this case Phytophthora agathidicida), and it follows the typical pattern of an introduced Phytophthora: a non-native strain of the disease, to which the local trees have very little resistance, is typically carried into the forest on the soles of hikers' shoes, where it then infects the root system, causing rapid dieback and mortality. Once established in an area, Phytophthora is very difficult to control; the primary solution is to prevent infection by excluding humans from the area. Alternatively, visitors can be required to follow a few simple but hard-to-enforce control measures (basically, clean your shoes after each hike and spray them with a chlorine bleach solution right before entering the sensitive area). As of late 2017 kauri dieback has been reported in many of the most popular kauri areas, including Waitakere Ranges Regional Park, and of Conservation reserves at Okura, Albany, Pakiri, Great Barrier, the Coromandel Peninsula, Trounson Kauri Park and Waipoua Forest (Morton 2017). For further information, visit the Kauri Dieback site. It is also interesting that dogs have been trained to sniff out Phytophthora-infected trees (de Jong 2021).
Remarkable Specimens
The kauri is the most famous native tree in New Zealand, largely because it is among the largest trees in the world. Tane Mahuta (Maori for 'Lord of the Forest'), in Waipoua Forest Sanctuary, pictured here, is commonly claimed to be the largest. In 2001, it was measured at 45.2 m tall with a dbh of 491 cm; that number is misleading, though, because the base of the tree is buttressed. The main trunk is 17.8 m tall and contains 255 m3 of wood. The total wood volume, including the crown, is much larger. The enormous crown is nearly 35 m wide and has a dozen branches over 1 m in diameter, three greater than 2 m, and one over 3 m. Adding the wood in the crown brings the total wood volume up to 516.7 m3, making this tree just a bit larger than the largest known Thuja plicata. Thus Agathis australis is the largest tree in the Araucariaceae and the third largest conifer, after Sequoiadendron and Sequoia (Robert Van Pelt e-mail, 2003.01.27). A sign posted at the tree by the Department of Conservation gives older, somewhat different numbers. The nearby Te Matua Ngahere is 16.4 m in girth, but only 29.9 m tall, giving it a stem volume of 208.1 m3. Many other specimens attain girths of more than 10 m (Burstall and Sale 1984 provide these figures and those of 18 other big trees as well). Sadly, much larger trees were cut during the orgy of logging that deforested most of the species' range. One in the Coromandel Peninsula was said to be 23.2 m in girth, and 24.4 m to the first limb. Such numbers are always questionable, but at least one early giant, Kairaru in the Tutamoe forest, was reliably measured at 20.1 m in girth with 30.5 m to the lowest limb and was revered by some as New Zealand's largest tree. Still, it fell to the saw in about 1900 (Adams 1986). The remaining giant kauris survived largely by being located in particularly dense and inhospitable bush; indeed, so impenetrable are these forests that some trees with a diameter of over 300 cm almost certainly remain to be found, most likely in Waipoua, Puketi or Omahuta Forests in Northland.
The tallest kauri on record, measured 1978, was 56.3 m, and grew in the upper Tairua River watershed; this tree is not visible in aerial photographs since 2016 and is presumed dead. Recent LIDAR data suggest the possibility of trees nearly 60 m tall in the Manaia Sanctuary, but there are so far no field measurements of these trees; the tallest measured specimen is a 53 m tree in Omahuta (Matt Smillie Facebook post in the "Great Trees of New Zealand" group, 2023.07.26).
The oldest known specimen is a subfossil tree documented in a tree-ring chronology covering the period 11175-9725 BCE, collected at Towai Farm in New Zealand by J. G. Palmer, P. Fenwick, and C. Turney (doi.org/10.25921/evvd-an62). The oldest tree in the study provided a 1397-year record. For trees in the historical era, the greatest demonstrable age I have found is from a log at The Kauri Museum that has been counted to have approximately 800 rings. The dendrochronological studies cited below have used considerably younger trees; the longest study I have seen gives a record that begins in 1580. As best I can determine, all very large trees that have died recently have had extensive internal rot, so precise ages for the largest trees may not be determinable. Ahmed and Ogden (1987) cored a variety of trees and inferred ages by extrapolation (a questionable procedure, but it can be useful), and "concluded that the 'normally attainable age' is >600 years" and that "individuals >2m d.b.h. probably often exceed 1000 years, but there is no reliable evidence for trees >2000 years in age."
Ethnobotany
Human use of the kauri began with exploitation by the native Maori people of New Zealand. "To the ancient Maoris the kauri ranked second only to the totara (Podocarpus totara) in importance. Some of the greatest northern war canoes were constructed out of single massive kauri trunks, felled in the forest after elaborate tapu ceremonies, then hollowed out with the use of fire and stone tools. The hulls of canoes made from single trunks in this way were known to reach 25 m. Burning kauri gum was used for heating and lighting, and a powder from the soot was used in tattooing" (Salmon 1996).
The following is some more information on the use of kauri to build large canoes (from a sign at Waitangi Treaty Grounds): "For Northern Maori the kauri was the preferred tree from which to build a waka taua [sea-going canoe]. Totara [Podocarpus totara] was more commonly used further south. Once a suitable tree was selected, it was blessed by a Maori tohunga (spiritual leader) who offered supplications to Tane Mahuta, God of the Forest. These rituals were also followed when a tree was cut and removed from the forest. Vegetation around a tree was cleared as a sign it had been earmarked for use. A tree so marked could be left standing for many years, allowing nature, Tane and Tawhirimatea (God of Winds) to step in and help the process. That natural process saw heavier timber develop on the tree's windward side, which then became the canoe's hull: strong, robust and stable in the water. In turn, lighter wood on the tree's leeward side made it easier to hollow into shape with adzes. Sometimes, to give nature a hand, the bark was peeled off one side of the tree, causing its wood to decay and making the task of hollowing out the trunk much easier. Sometimes fire was used to help fell and hollow out the tree but more often simple stone adzes were the only tools used. Once felled, levers were used to roll and turn the large logs, which could be up to two metres thick and 25 metres long." The canoe shown, possibly the largest ever built, was carved using similar techniques, except that steel tools were used, and the logs were hauled from the forest by a team of bullocks. It was carved from two giant kauris harvested in the Puketi Forest, and is capable of carrying 180 warriors.
From the Auckland Museum:
Northern Maori believed the kauri to be the father of the sperm whale. Because of their huge size, both are regarded as rangatira (chiefs) of their respective realms. Moreover, their bark and skin show similiarities of texture, while kauri gum is like the ambergris found in the intestines of the whale.
In times long past, a sperm whale came ashore and spoke thus to the kauri: "Kauri! Come with me to the sea which is fresh and cool."
"No!" said the kauri. "You may like the sea but I prefer to stand here with my feet in the soil."
"All right" said the whale "then let us agree to exchange our skins."
So that is why the bark of the kauri is thin and full of resinous oil.
To European settlers, kauri was an important timber source. The golden-colored wood, which appears iridescent when properly finished, is light, strong, and easily worked. Thanks to its high resin content it is resistant to decay, and because of the kauri's self-pruning ability, the wood is largely free of knots. It was greatly prized for ship masts and spars and for numerous other shipbuilding applications, and was the preferred timber for framing, finish carpentry, and cabinetmaking in its range. It was also used for railway carriages and road paving (Salmon 1996). The history of the kauri logging period has been much preserved; it is detailed by Adams (1986) and others, and the material culture of the day is splendidly preserved and described at The Kauri Museum. The gum of the kauri, also called copal, was used "for the manufacture of high quality paints and varnishes. It was obtained by digging over areas where kauri forests had been felled, and later by 'bleeding' kauri trees" (Salmon 1996). This exploitation of modern and subfossil deposits by 'gum diggers' caused the excavation of substantial tracts in the Northland. Fragments of this resin were gathered and polished with rather lovely results (see photos at the website for The Kauri Museum).
The surviving kauri forests are fully protected from logging, and so far there has been little effort to use kauri (or, indeed, any other native trees) in commercial forest plantings in New Zealand. Current exploitation is thus focused on the subfossil remnants of past forests preserved in anaerobic sediments, locally referred to as "swamp kauri." Most of this wood is used by craftsmen to produce bowls, fine furniture and the like; the subfossil deposits also yield copal used for jewelry and related arts.
Note: Kirk's Forest Flora (1889) is said to have accounts of the contemporary uses of kauri (Allan 1961).
The species is dendrochronologically usable. The earliest work is described by Bell and Bell (1958) and more early work is recounted by Ahmed and Ogden (1985). Research up to about 1992 is summarized by Palmer, Ogden and Norton (Norton & Palmer 1992, Palmer & Ogden 1992) and has included population age structure work as well as climate reconstruction (e.g., Burrows & Greenland 1979, Salinger et al. 1994). A synthesis of these studies was recently published by Fowler et al. (2004). The most detailed dendroclimatic work to date was completed by Buckley et al. (2000) and Fowler et al. (2000) and uses a network of nine tree-ring chronologies to explore the climate-growth relationship and show an ENSO (El Niño-Southern Oscillation) signal in the data.
A curious phenomenon with this species is the occurrence of extensive subfossil deposits, currently being extensively exploited for craftwood. These deposits, often including whole logs over 2 m in diameter, have been radiocarbon dated to have ages of from 3,800 to over 40,000 years, raising the possibility of developing exceedingly long chronologies from subfossil material (but see Remarks).
From the Kauri Museum:
There is an extensive patchwork of buried kauri sites found in Northland, especially in coastal regions behind dunes. ... The buried logs are being 'harvested' for commercial purposes because the wood is perfectly preserved. Some of the buried trees are enormous, with diameters greater than 3 m. This wood and associated preserved material, represents an immense store of information about past environments, with some buried wood being radiocarbon dated more than 50,000 years old.
Observations
The kauri forests have been so much reduced by logging that there are relatively few places left where a person can view native kauri groves. On the Coromandel Peninsula, a small grove is associated with the Square Kauri, prominently signed and visible from the road about 8 km east of Tapu on the road to Coroglen. This is the largest surviving kauri outside of Northland. Another fine grove, requiring a walk of several hours, can be seen up the Kaueranga Valley east of Thames. Check at the Department of Conservation visitor center about 15 km east of Thames for maps and more information. This is also a good place to see kauri dams, temporary dams that were opened up to flush kauri logs downstream to the mills during the early days of kauri logging. There is also a kauri grove in the north Coromandel, about 6 km south of Coromandel town; I have no more information about it, though.
There are kauri on Great Barrier Island, but the island was logged and the kauri there represent natural regeneration. The area has several well-preserved kauri dams.
Most of the other surviving kauri are found from Auckland northwards. Some fine groves are preserved in the Waitakere Ranges Regional Park just west of Auckland, with some of the best walks visible along the Cascades Walk, a short (ca. 1 hour) tramp accessed from the Waitakere Golf Course. Here you can see both uncut native forest, and regeneration forest that dates from logging in about 1920.
Farther north, Trounson Kauri Park, 7 km off Route 12 from Dargaville to Waipoua Forest, provides a memorable kauri experience that should not be missed. The short (ca. 1 hour) walk passes through an old kauri grove with some trees over 300 cm diameter. Younger trees are coming up in the understory, providing graphic illustration of how gap-phase regeneration can produce a perpetuating kauri forest. The area has been very intensively controlled for exotic invasive predators (possums and mustelids, the bane of New Zealand wildlife), and thus affords an exceptionally pristine wildlife and flora as well.
The largest kauris, the largest area of intact kauri forest, and the greatest variety of walks are to be found in the Waipoua Forest. Short (10 minutes to 1 hour) walks take you to Tane Mahuta and Te Matua Ngahere, the two largest known kauris. Walks of up to three days are possible on other tracks in the park. See also the website, Waipoua Forest.
Finally, the northernmost large kauri stands are to be found in the Puketi and Omahuta Forests just northwest of Kaikohe. Each forest has a short walk (Omahuta Kauri Sanctuary Walk and Puketi Nature Trail) that will get you into an impressive kauri grove. These forests also contain one of the largest continuous tracts of native forest left in Kauri country and have few tracks or trails; thus the area is potentially the most significant for discovering new giant kauris.
Remarks
"Abundant sub-fossil wood of kauri Agathis australis (Salisb.) is preserved in peat swamps throughout the present distribution of the species in North Island, New Zealand. Analysis of 107 radiocarbon dates on this material shows they fall mainly into two groups. A northern group represents late Pleistocene interstadial forests, while a southern group relates to a mid to late Holocene expansion to the current southern limits of the species at c. 38°S. Wood growth rates in the interstadial samples, compared to modern trees, suggest that kauri was stressed by lower temperatures and a wetter environment. With further temperature reduction in the late Otiran stadial (last glacial maximum) growth may have been limited to c. four summer months. The scarcity of kauri pollen from stratigraphic samples and the rarity of wood dating from this period support the contention that kauri was much reduced in abundance during the last glacial maximum" (Ogden et al. 1992).
Subfossil kauri has also been used to study variation in the earth's magnetic field strength, which is a major determinant of the rate at which radiocarbon is generated in the upper atmosphere. In 2019 a subfossil tree was excavated that had lived between 42,500 and 41,000 years ago, and recorded in its rings a partial collapse of the Earth's magnetic field (Osborne 2019).
Young kauri trees are called 'rickers'. In the mid-19th century, the timber blanks for masts and spars were called rickers by British seamen, a corruption of 'Riga', the Baltic port that traditionally had been the chief export port for such timbers. The kauri were put to the same use shortly after the British colonization of New Zealand (pers. comm. Graeme Platt 2003.03.12).
Citations
Adams, J. G. Erne. 1986. Kauri: a king among kings. 2nd rev. ed. Auckland: Wilson & Horton Ltd. ISBN 0-86864-080-8.
Ahmed, M. and J. Ogden. 1985. Modern New Zealand tree-ring chronologies III. Agathis australis Salisb. kauri. Tree-Ring Bulletin 45:11-24.
Ahmed, M. and J. Ogden. 1987. Population dynamics of the emergent conifer Agathis australis (D. Don) Lindl. (kauri) in New Zealand. I. Population structures and tree growth rates in mature stands. New Zealand Journal of Botany 25(2):217-229.
Allan, H. H. 1961. Flora of New Zealand. Volume I, Indigenous Tracheophyta. Wellington: R.E. Owen Government Printer.
Bell, V., and R. E. Bell. 1958. Dendrochronological studies in New Zealand. Tree-Ring Bulletin 22:7-11.
Buckley, B., J. Ogden, J. Palmer, A. Fowler, and J. Salinger. 2000. Dendroclimatic interpretation of tree-rings in Agathis australis (kauri). 1. Climate correlation functions and master chronology. Journal of the Royal Society of New Zealand 30(3):263-276.
Burns, B.R. and J. R. Leathwick. 1996. Vegetation-environment relationships at Waipoua Forest, Northland, New Zealand. New Zealand Journal of Botany 34: 79-92.
Fowler, A., J. Palmer, J. Salinger, and J. Ogden. 2000. Dendroclimatic interpretation of tree-rings in Agathis australis (kauri). 2. Evidence of a significant relationship with ENSO. Journal of the Royal Society of New Zealand 30(3):277-292.
Fowler, A., G. Boswijk, and J. Ogden. 2004. Tree-ring studies on Agathis australis (kauri): A synthesis of development work on late Holocene chronologies. Tree-Ring Research 60(1):15-29.
Loudon, J. C. 1829. An Encyclopaedia of Plants, p. 802. Available: Biodiversity Heritage Library, accessed 2023.03.04.
Morton, Jamie. 2017.11.30. Council mulls mulls Waitakeres closure over kauri crisis. New Zealand Herald. Available: http://www.nzherald.co.nz/nz/news/article.cfm?c_id=1&objectid=11949906, accessed 2017.12.01.
Osborne, Hannah. 2019.07.04. Ancient Tree With Record of Earth's Magnetic Field Reversal in Its Rings Discovered. Newsweek online.
See also
Ahmed, M. 1984. Ecological and dendrochronological studies on Agathis australis Salisb. (Kauri). Ph.D. dissertation, Auckland University, New Zealand.
Bridge, M. C. and J. Ogden. 1986. A sub-fossil kauri (Agathis australis ) tree-ring chronology. Journal of the Royal Society of New Zealand 16(1):17-23.
Collins, L. and B. Burns. 2001. The dynamics of Agathis australis-Nothofagus truncata forest in the Hapuakohe Ecological District, Waikato Region, New Zealand. New Zealand Journal of Botany 39:423-433.
Ecroyd, C. E. 1982. Biological flora of New Zealand 8. Agathis australis (D. Don) Lindl. (Araucariaceae) Kauri. New Zealand Journal of Botany 20:17-36. ABSTRACT: Information relevant to the ecology of Agathis australis is presented, together with a distribution map based on its presence or absence in grid squares and bibliographic references to other information.
Enright, N. J., R. M. Bartlett and C. R. De-Freitas. 1993. Patterns of species composition, recruitment, and growth within canopy gaps in two New Zealand
kauri (Agathis australis) forests. New Zealand Journal of Botany 31(4):361-373.
Forest Phytophthoras of the World provides substantial information, educational materials, and a detailed bibliography detailing the Phytophthora infection of kauri.
Fowler, A. 1984. A dendrochronological study of kauri (Agathis australis). M.Sc. thesis, Auckland University, New Zealand.
Lorrey, A. M., G. Boswijk, A. Hogg, J. G. Palmer, C. S. M. Turney, A. M. Fowler, J. Ogden, and J.-M. Woolley. 2018. The scientific value and potential of New Zealand swamp kauri. Quaternary Science Reviews 183:124–139.
McKenzie, E. H. C., P. K. Buchanan, and P. R. Johnston. 2002. Checklist of fungi on kauri (Agathis australis) in New Zealand. New Zealand Journal of Botany 40:269-296.
Mitchell, N. D. 1991. The derivation of climate surfaces for New Zealand, and their application to the bioclimatic analysis of the distribution of kauri (Agathis australis). Journal of the Royal Society of New Zealand 21(1):13-24.
The New Zealand Plant Conservation Network, accessed 2010.11.22.
Norton, D. A., J. G. Palmer and J. Ogden. 1987. Dendroecological studies in New Zealand 1. An evaluation of tree age estimates based on increment cores. New Zealand Journal of Botany 25: 373-383.
Ogden, J., A. T. Wilson, C. H. Hendy, A. G. Hogg, and R. M. Newnham. 1992. The late Quaternary history of Kauri (Agathis australis) in New Zealand and its climatic significance. Journal of Biogeography 19:611-622.
Ogden, J., G. M. Wardle and M. Ahmed. 1987. Population dynamics of the emergent conifer Agathis australis (D. Don) Lindl. (kauri) in New Zealand. II. Seedling population sizes and gap-phase regeneration. New Zealand Journal of Botany 25(2):231-242.
Owens, John N., Glenda L. Catalano and Jenny Aitken Christie. 1997. The reproductive biology of kauri (Agathis australis). IV. Late embryogeny,
histochemistry, cone and seed morphology. International journal of plant sciences 158(4):395. (Parts I, II, III also published).
Sale, E. V. Quest for the kauri : forest giants and where to find them. ASIN: 0589011235
Silvester, W. B. and T. A. Orchard. 1999. The biology of kauri (Agathis australis) in New Zealand II. Nitrogen cycling in four kauri forest remnants. New Zealand Journal of Botany 37.
Silvester, W. B. 2000. The biology of kauri (Agathis australis) in New Zealand I. Production, biomass, carbon storage, and litter fall in four forest remnants. New Zealand Journal of Botany 38:205-220.
Stephens, D. W., W. B. Silvester, and B. R. Burns. 1999. Differences in water-use efficiency between Agathis australis and Dacrycarpus dacrydioides are genetically, not environmentally, determined. New Zealand Journal of Botany 37.
Turney, C. S. M., L. K. Fifield, A. G. Hogg, J. G. Palmer, K. Hughen, M. G. L. Baillie, R. Galbraith, et al. 2010. The potential of New Zealand kauri (Agathis australis) for testing the synchronicity of abrupt climate change during the Last Glacial Interval (60,000–11,700 years ago). Quaternary Science Reviews 29(27):3677–3682.
Gymnosperms of New Zealand.