Pinus pseudostrobus
Lindley 1839
Common names
Macochyaj, macohtai, pino blanco, pino lacio, pino liso, pino real (Lopez-Upton 2003). The scientific name, pseudostrobus, literally means a false Pinus strobus. P. strobus is a white pine native to the eastern U.S. and Canada, and it would be an inattentive botanist indeed who could confuse the two species; neither trees, nor foliage, nor cones bear the slightest resemblance to each other.
Taxonomic notes
The taxonomy of this species has been controversial, with very different perspectives presented in two monographs addressing the problem, those of Perry (1991) and of Farjon and Styles (1997). The basic problem is that of Mexican pines in general, i.e., that Mexico is the center of diversity for the genus Pinus and appears to be one of the few remaining places where new conifer species are actively arising. P. pseudostrobus sensu latu is a very diverse taxon with a wide geographical and environmental range. The species can hybridize in nature with its close relative P. montezumae (Perry 1991 cited in Lopez-Upton 2003). To complicate matters further, there is no type specimen, nor even a good description of the type. In a situation like this, there is no firm objective basis for definition of the species. In this case more than most, the "species" we name is little more than a tag for a large and diverse collection of populations that are distinguishable from other, closely related taxa on the basis of (relatively) easily observed characters. They may nonetheless contain, collectively, a great deal of genetic variability. A really thorough evaluation of relationships within the taxon would require evaluation of populations from throughout its range. Farjon and Styles (1997) worked toward that goal primarily by systematic comparison of a large number of herbarium specimens, supplemented by some field observation and detailed review of the literature. Perry (1991), on the other hand, worked as a field biologist with very extensive experience throughout the range of the species, and a deep familiarity with the ecological and silvicultural literature. I do not question that Perry observed the differences that he saw; however, his level of taxonomic rigor was less than that of Farjon and Styles. By Farjon's account, and by the keys that Perry presents, it appears that Perry would sometimes discriminate species according to inconsistent morphological characters. My own fieldwork, though much less extensive than that of the monographers, has not refuted Farjon's highly parsimonious taxonomic approach. Accordingly the classification presented here is that of Farjon and Styles (1997). Since then, this species has also been addressed in molecular studies that have tended to confirm the systematics of Farjon and Styles (1997); see the "Taxonomic notes" for Pinus ponderosa for further discussion of those studies.
In this treatment I recognize three infraspecific taxa:
- P. pseudostrobus var. pseudostrobus forma pseudostrobus (type, Michoacán: near Angangueo, 1838, Hartweg s.n.)
- P. pseudostrobus var. pseudostrobus forma protuberans Martínez 1948 (type, Distrito Federal: Eslava, 1940, M. Martínez 3431)
- P. pseudostrobus var. apulcensis (Lindley) G.R. Shaw 1909
Of these, the type forma is by far the most widely distributed and common one, and is described in detail on this page. Forma protuberans is relatively uncommon and sparsely distributed and I have found little information on it; it is mentioned below as applicable. Var. apulcensis is clearly distinct and has been treated by several respected authors as a distinct species (P. apulcensis or P. oaxacana), and for those reasons is treated on a separate page.
Synonyms for the type variety, forma pseudostrobus (type, Michoacán, near Angangueo, 1838, Hartweg s.n.) (Farjon and Styles 1997, Farjon 1998):
- P. orizabae G. Gordon 1846 (type, Pico de Orizaba, 1838?, Hartweg s.n.)
- P. protuberans Roezl 1857 (type, México: Contreras, 1857, Roezl s.n.)
- P. angulata Roezl 1857
- P. regeliana Roezl 1857 (type, Volcán Iztaccihuatl, on SW side of volcano, near Hacienda de Zavaleta, 1857, Roezl s.n.).
- P. protuberans Roezl var. angulata (Roezl) Carrière 1867
- P. pseudostrobus Lindley var. estvezii Martínez 1945
- P. pseudostrobus Lindley var. coatepecensis Martinez 1945 (type, Veracruz: "Cerea de Coatepec," 1940, M. Martínez 3432)
- P. pseudostrobus Lindley var. apulcensis (Lindley) Martínez, 1945 non Shaw 1909
- P. pseudostrobus Lindley forma megacarpa Loock. 1950 (type, Michoacán: Ciudad Hidalgo, Loock 10618)
- P. estevezii (Martínez) J. P. Perry 1982 (type, Nuevo León: Cañon de Mieleras, 1941, M. Martínez 3433)
- P. pseudostrobus Lindley subsp. apulcensis (Lindley) sensu Stead, excl. basion. & type, 1984 (type, Hidalgo: Apulco, 1941, M. Martínez 3434)
- P. nubicola J. P. Perry 1987 (type, Guatemala: 40 km E of San Jose Pinula along rd. to Mataquescuintla, 1979, Perry GUA-32-79).
- P. pseudostrobus Lindley var. laubenfelsii Silba 1990 (type, Sinaloa: mtns. E of Mazatlan, 1968, Laubenfels 663)
Description
Evergreen trees 20-40(-45) m tall and up to 100 cm DBH with a round trunk and monopodial architecture with long, spreading and ascending, usually whorled first-order branches; higher-order branches slender, spreading, downcurved to ascending. Crown depth normally 50-67% of tree height. Bark on young trees smooth, red-brown to grey-brown, with age becoming darker, thick and scaly, with elongated plates and deep longitudinal fissures. Shoots slender, smooth, glaucous or pruinose, with short decurrent pulvini. Cataphylls 10-15 mm long, soon recurved, with ciliate margins, reddish brown to dark brown, falling shortly after the fascicles. Fascicle sheaths (15-)20-30(-35) mm long, persistent, red-brown, weathering grey-brown. Leaves in fascicles of 5, rarely 4 or 6, in lax tufts at the ends of upturned branches, persisting 2-3 years, slender, straight, spreading or drooping, (18-)20-30(-35) cm × 0.8-1.3 mm, with serrulate margins, glaucous-green. Stomata on all faces of leaves, in (2-)3-7 lines on the outer face and 2-4(-5) lines on each inner face. Pollen cones crowded near the basal end of a new shoot, numerous, spreading, ovoid-oblong to cylindrical, 20-35 × 5-7 mm, yellow, turning brown. Seed cones subterminal, solitary or in pairs (rarely whorls of 3-4) on very short, stout peduncles which remain with a few cone scales on the branch when the cone has fallen. Immature cones ovoid, 15-20 × 10-12 mm, with small, spreading spines, purple-brown, maturing in two seasons. Mature cones ovoid-oblong, asymmetrical, often curved at base, ovoid-oblong to broad-ovoid when opened, then 7-16 × 6-13 cm. Seed scales ca. 140-190, parting to release the seeds except the proximal infertile scales, usually thick woody, oblong, straight or slightly curved, red-brown to dark purple-brown. Apophysis extremely variable, from nearly flat to elongated, more so on one side of the cone and toward the base, transversely keeled, tapering to an obtuse or mucronate umbo, rhombic or pentagonal, upper margin angular, irregularly undulate, or rounded, colour in various hues of brown. Umbo dorsal, variable, from obtuse to prominent, 3-15 mm long, 5-10 mm wide at base, without a prickle or prickle deciduous, usually darker than the apophysis. Seeds obliquely ovoid, slightly flattened, 5-7 × 3-4.5 mm, ochraceous to grey-brown, with or without dark spots. Seed wings articulate, effective, held to the seed by two claws, thinly covering part of the seed on one side, obliquely-ovate, with a straight side, 20-25 × 7-10 mm, yellowish brown, translucent, distal portions darker. Seedlings: The hypocotyl of this species is consistently longer (mean 31.8 mm) than that of P montezumae (mean 17.5 mm), according to a study by Caballero (1967). Cotyledons (7-)8-l2(-13), but given as a mean of 7.2 by Caballero (1967). Pollen dispersal occurs in February-April, depending on location and altitude (Farjon and Styles 1997).
The species begins reproducing at 6 to 7 years and flowers in February and March (Patiño-Valera 1973, cited in Lopez-Upton 2003). The interval between large cone crops is from 3 to 5 years (Zamora-Serrano and others 1993, cited in Lopez-Upton 2003). In central Mexico, seeds average 53,705 per kg. In Chiapas, México, seeds average 44,500 per kg (Patiño-Valera 1973, Zamora-Serrano and others 1993, both cited in Lopez-Upton 2003). Wood production is 12 to 30 m3/ha/yr (Pancel 1993, cited in Lopez-Upton 2003).
Farjon and Styles (1997) provide the following key to the infraspecific taxa:
1. |
Seed scales with slightly to prominently raised or elongated apophyses; umbo obtuse or elongated, more or less continuing the line of the transverse keel on the apophysis. |
2. |
1. |
Seed scales with nearly flat apophyses; umbo acutish, with a distinct angle to the transverse keel of the apophysis. |
var. pseudostrobus forma protuberans |
2. |
Seed scales with slightly to prominently raised but not elongated apophyses; umbo not elongated, obtuse. |
var. pseudostrobus forma pseudostrobus |
2. |
Seed scales with prominently raised and, at least in the lower half of one side of the cone, elongated apophyses; umbo on these scales also elongated. |
var. apulcensis |
Distribution and Ecology
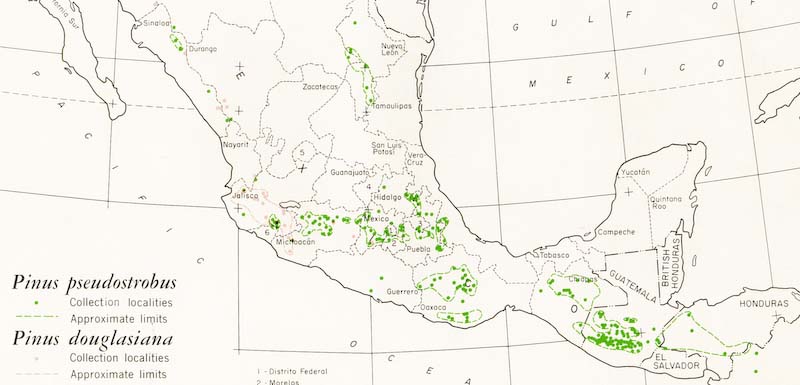
For the species as a whole: N El Salvador, Guatemala (highlands), W Honduras, Mexico: Sinaloa-Durango border, Nuevo León, SE Coahuila, E Guanajuato(?), Jalisco, Michoacán, México, Distrito Federal, Morelos, Hidalgo, Puebla, Tlaxcala, W-C Veracruz, Guerrero, Oaxaca, Chiapas. A 250 km range gap at the Isthmus of Tehuantepec separates the Mesoamerican populations from the Mexican populations. Other disjunctions of comparable size separate the NW (Sinaloa-Durango) and NE (Nuevo León-Coahuila) populations from the main distribution in central Mexico (Farjon and Styles 1997).
Zone 9 (cold hardiness limit between -6.6°C and -1.1°C) (Bannister and Neuner 2001).
Distribution data from USGS (1999). Points represent isolated or approximate locations.
Pinus pseudostrobus forms pure stands or grows in association with P. montezumae, P. douglasiana, P. devoniana, P. maximinoi, P. leiophylla, P. ayacahuite, P. patula, P. cembroides, P. hartwegii, P. pringlei, Abies religiosa, Quercus sp., Arbutus sp., Juniperus sp., Buddleia sp., and Dasylirion sp. (Eguiluz-Piedra 1978, Perry 1991, both cited in Lopez-Upton 2003).
The best stands are found at 2500 m on deep volcanic soils. This tree can also be found in swallow and calcareous soils. Pinus pseudostrobus grows in temperate to temperate-warmer climates, where temperatures may drop to freezing during the coldest winter months. The species is found where temperatures range from -9 to 40°C and annual rainfall from May to October is 600 to 2000 mm (Eguiluz-Piedra 1978, Martínez 1948, Perry 1991, both cited in Lopez-Upton 2003).
In frequent-fire environments the seedlings may express a "grass stage." See the "Remarks" section HERE for details on "grass stage" growth.
Forma pseudostrobus is the most widespread variety, occurring throughout the entire range of the species. The other two varieties are often sympatric. Forma pseudostrobus is in many places a common or abundant pine of montane to high montane habitat in the cold-temperate to warm-temperate zones. Its altitudinal range varies little along the entire 2500 km of its range: (850-)1900-2400(-3250) m. Annual precipitation is at a minimum of ca. 800 mm, but in Guatemala and Honduras it can be more than 2000 mm. The taxon is often dominant in mixed conifer, pine, and pine-oak forests, occasionally associated with Liquidambar as on the wet Gulf Coast side of the mountains of central and southern Mexico. In its driest habitat in central and northern Mexico it occurs with Pinus cembroides, Juniperus flaccida, and Quercus spp., with an understorey of, e.g., Agave, Buddleja, Opuntia, and Salvia (Farjon and Styles 1997).
Forma protuberans is recognized by Farjon only from the state of Mexico and in the Ajusco Mountains near Mexico City. Farjon, however, acknowledges that Martínez (1948) and Perry (1991) also recognize it from Mexico: Distrito Federal, S Jalisco, México, Michoacán, Morelos, Puebla, and one locality in Oaxaca.
Remarkable Specimens
A specimen near Magoebaskloof in the Woodbush Forest Reserve, South Africa, was measured on 2011.08.15 as being 50.3 m tall and 136 cm dbh (Leon Visser email 2014.11.05). This is one of a group of 3 P. pseudostrobus growing in an area of native forest.
Ethnobotany
This is one of the most commonly harvested timber pines in southern Mexico and the highlands of Guatemala and parts of Honduras. Its light, soft, strong, pale yellow wood has a specific gravity of 0.32 to 0.51 and is only slightly resinous. It produces high pulp yields and is relatively free of knots, thus widely used for general construction, hewn timber, and decorative items, as well as firewood. Exploitation for timber has led to regional depletion of the best stands. In Mexico, it is also used as a source of resin (Farjon and Styles 1997; and Eguiluz-Piedra 1978, Perry 1991, Wright and Malan 1991, Wright and Wessels 1992, Zobel 1965, all cited in Lopez-Upton 2003).
The species has been used in dendrochronological research. One study performed in Oaxaca (Naylor 1971) found generally unsatisfactory results: all sampled trees were young, complacent, and could not be crossdated. This finding was attributed to an insufficiently seasonal climate. I am cautious about this, knowing that Naylor (1971) reached the same conclusion for Pinus hartwegii, a species which has since proved useful not in Oaxaca, but at similar sites in Colima.
Observations
I saw it forming nearly closed-canopy forests with many trees over 30 m tall and 100 cm DBH, in forests east of Cortes Pass, which lies beteen Popocatepetl and Ixtaccihuatl, where it grows extensively with P. hartwegii and, somewhat lower, with P. montezumae and P. strobiformis subsp. veitchii. I suspect that hybrids with P. montezumae occur in this area as well; certainly there were many trees that we could not confidently assign to one or the other species.
Remarks
The epithet pseudostrobus seems to refer to a perceived resemblance to Pinus strobus (Farjon 2010), although the two share no characters apart from the fact that both are pines.
This species is a principal host for the dwarf mistletoes Arceuthobium aureum subsp. aureum (in Guatemala), A. aureum subsp. petersonii (in Oaxaca and Chiapas), A. durangense (in Jalisco), A. globosum subsp. grandicaule (in southern Mexico and Guatemala), and A. oaxacanum (rare, in Oaxaca) (Hawksworth and Wiens 1996).
Leptoglossus occidentalis and Conophthorus ponderosae are the most important insect pests of conelets, cones, and seeds. Tetyra bipunctata produces empty seeds. Cecidomyia bisetosa causes cone death. Larvae of Cydia montezuma and Megastigmus albifrons feed on seeds (Cibrián-Tovar and others 1995, cited in Lopez-Upton 2003). Seeds and saplings are severely affected by the fungus Shaeropsis sapinea (Fr.) (Rees and Webber 1988, cited in Lopez-Upton 2003).
Citations
Lindley, J. 1839. Edwards's Botanical Register 25:62.
López-Upton, Javier. 2003. Pinus pseudostrobus. Species description in the Tropical Tree Seed Manual. Available http://www.rngr.net/Publications/ttsm/Folder.2003-07-11.4726 (accessed 2007.08.31).
Naylor, T.H. 1971. Dendrochronology in Oaxaca, Mexico: a preliminary study. Tree-Ring Bulletin 31:25-29.
See also
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library. This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
Stead and Styles 1984.