(both subspecies.)
Pinus leiophylla
Schiede ex Schlechtendal et Chamisso 1831
Common names
Ocote, ocote prieto, pino chino, smooth-leaved pine (Perry 1991, Martínez 1948, both cited by López-Upton 2003).
Taxonomic notes
Two subspecies, P. leiophylla subsp. leiophylla and P. leiophylla subsp. chihuahuana (Engelm.) Shaw 1909. Subsp. chihuahuana was separately described by Engelmann in Wislizenius 1848 as Pinus chihuahuana, and the similarities between that taxon and Pinus leiophylla were discerned by Shaw (1909). Some later authors continue to discriminate the two species, but recent genetic analyses using both nuclear and plastid markers have generally confirmed a sister relationship between the taxa, while placing them both very close to P. lumholtzii (Gernandt et al. 2018, Cruz-Nicolas et al. 2024). One phylogeographic study suggests a Pleistocene divergence for the two taxa (Rodríguez‐Banderas et al. 2009), which, combined with differences in their distributions and ecological settings (discussed below), supports distinguishing the taxa at subspecies rank.
Typical of early-collected pines (this species was described in 1831, along with Pinus patula), the holotype consists of a single sheet with two sterile branchlets.
Description
Tree, 15-35 m tall and 20-85 cm dbh, usually with a single round trunk often clear of branches for 60% of height, and an open, rounded crown. Bark dark grey-brown, very thick, rough, scaly, divided into elongate irregular scaly plates with deep longitudinal fissures; on young trees and branches scaly, flaking, red-brown to grey-brown. Branches long, slender, spreading or ascending, the lower ones curved downwards. Shoots scaly with short, decurrent pulvini, red-brown or glaucous turning grey-brown. Fascicle sheaths 12-20 mm long with 5-6 early-deciduous scales, so that mature fascicles have no sheath. Leaves in fascicles of (2-)3-5(-6), spreading in lax or rigid tufts near the ends of branches, persisting 2-3 years, (4-)6-15(-17) cm × 0.5-1.3(-1.5) mm, margins minutely serrulate, light green, stomata on all faces in 4-8 lines on the convex abaxial face, 3-4 lines on the adaxial faces. Resin ducts 2-6, medial, sometimes 1-2 internal, stele round with 2 vascular bundles. Pollen cones crowded near end of shoots in a short dense cluster, cylindrical, 10-15(-20) × 4-6 mm, yellow-pink maturing light brown. Seed cones subterminal, solitary or in whorls of 2-5 on stout peduncles 10-20 mm long, persisting several years after opening. Immature cones ovoid to subglobose, pink turning purple-green to purple-brown, maturing in three seasons, falling 2-3 years after maturity. Mature cones ovoid, nearly symmetrical, (4-)5-7(-8) × (3-)4-5.5 cm when open. Seed scales 50-70, straight or recurved near base of cone, purple-brown with lighter grey marks of seed wings. Apophysis raised, transversely keeled, with the second season's growth showing as a narrow band around the umbo; rhombic, up to 15 mm wide, brown. Umbo dorsal, pyramidal, with a blunt prickle, darker than the apophysis. Seeds ovoid, 3-4(-5) mm long, dark grey-brown with black spots. Seed wings articulate, oval, 10-18 × 4-8 mm, yellow-brown, translucent. Adventitious shoots may sprout after fire or cutting (Martínez 1948 cited by López-Upton 2003, Farjon and Styles 1997). The wood has a specific gravity of 0.44 to 0.51, and is relatively dense, heavy, hard, and very
resinous with a pale brown heartwood (Echenique-Manrique and Díaz-Gómez 1969, Murillo 1988, Zobel 1965, all cited by Lopez-Upton 2003).
Subsp. leiophylla has leaves chiefly in fascicles of 5, 0.5-0.9 mm wide, with 4-6 lines of stomata on the abaxial face and 2-3 resin ducts. Subsp. chihuahuana has leaves chiefly in fascicles of 3, 0.9-1.3 mm wide, with 5-8 lines of stomata on the abaxial face and 4-6 resin ducts (Farjon and Styles 1997).
The early-deciduous fascicle sheaths on a hard pine represent a simple, reliable character that makes it easy to identify this species in the field.
Distribution and Ecology
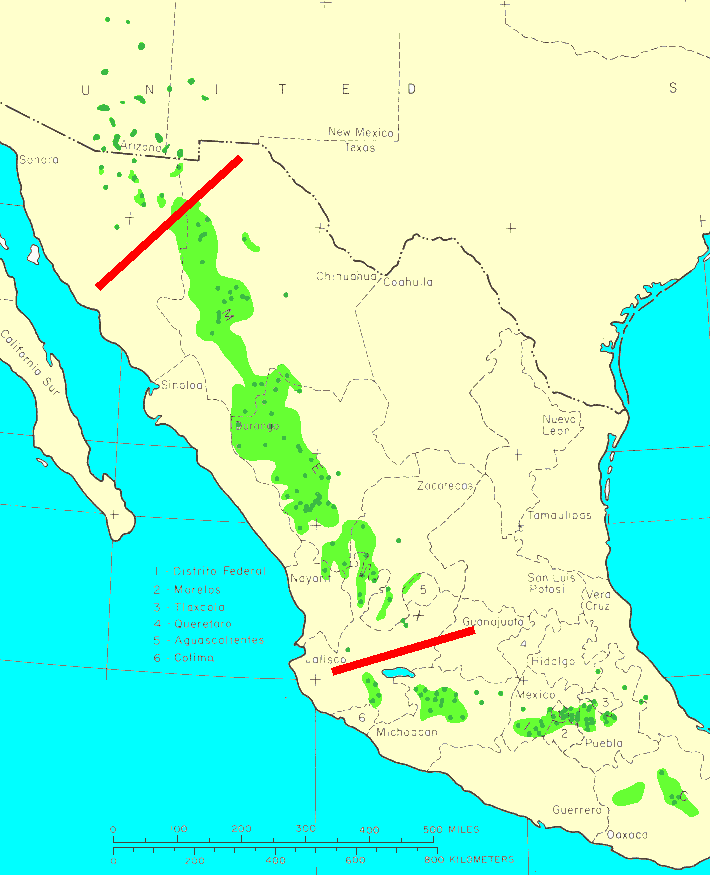
Mexico: Sonora, Chihuahua, Durango, Nayarit, Zacatecas, Jalisco, Michoacán, México, Distrito Federal, Hidalgo, Morelos, Tlaxcala, Puebla, Veracruz, Guerrero, and Oaxaca; USA: Arizona and New Mexico; at 1500-3300 m elevation. From Zacatecas northward subsp. leiophylla is gradually replaced by subsp. chihuahuana (Farjon and Styles 1997). As described above, in this overlap zone, subsp. leiophylla is primarily a tree of maritime/warm/mesic sites west of the Sierra Madre Occidental crest, while subsp. chihuahuana is characteristic of continental/cold/xeric sites east of the crest.
Bannister and Neuner (2001) say it is hardy to Zone 9 (cold hardiness limit between -6.6°C and -1.1°C), but they must only be considering subsp. leiophylla; subsp. chihuahuana is substantially more hardy.
Distribution data from USGS (1999).
Found at (1600)-2200-2750-(3000) m elevation, which corresponds to temperate to warm-temperate climates (Perry 1991 cited by López-Upton 2003) having an annual temperature range of -15 to 38°C and annual rainfall of 700 to 1500 mm with a May-October wet season (Eguiluz-Piedra 1978 cited by López-Upton 2003). It grows best on deep, well-drained soils of volcanic origin (Martínez 1948 cited by López-Upton 2003).
The species rarely forms pure stands, and is commonly associated with other pines including P. engelmannii, P. arizonica, P. teocote, P. lumholtzii, P. montezumae, P. ayacahuite, P. pseudostrobus (both subsp.), P. devoniana, and P. patula, as well as Abies religiosa, A. guatemalensis, Juniperus sp., Arbutus sp., Quercus sp., and several hardwoods (Eguiluz-Piedra 1978, Perry 1991, both cited by López-Upton 2003). See also Thompson et al. (1999).
Remarkable Specimens
I only have data for subsp. chihuahuana, which see.
Ethnobotany
I have no data on its aboriginal uses. Since the mid-20th Century, exploitation of this species has increased throughout much of its range, particularly in the Sierra Madre Occidental. It is tapped for resin and cut for general construction, railway ties, and fuelwood (Eguiluz-Piedra 1978, Perry 1991, both cited by López-Upton 2003; Farjon and Styles 1997). Subsp. chihuahuana is particularly exploited for resin production (Farjon and Styles 1997).
Observations
See the pages for each subspecies.
Remarks
Pinus leiophylla typically produces some cones every year, masting at intervals of 3 to 5 years (Zamora-Serrano et al. 1993, cited by López-Upton 2003). Cones are susceptible to the fungus Caeoma conigenum Heds. et Hunt (Martínez 1948, Perry 1991, both cited by López-Upton 2003). Important insect pests are Conophthorus conicolens Wood, which can destroy 16 to 60 percent of conelets, cones, and seeds, and Leptoglossus occidentalis
Heidemann (Cibrián-Tovar et al. 1995 cited by López-Upton 2003).
I puzzled about the differences between leiophylla and chihuahuana for several days as I criss-crossed the Sierra Madre Occidental, first heading east on Mex-16 from Hermosillo to Chihuahua and then back west on Mex-40 from Durango to Mazatlán, stopping in dozens of places to examine the two subspecies. There appears to be a continuum of intermediate form. The endpoints, taken alone, are quite different trees, and it's usually possible to assign a given tree to leiophylla or chihuahuana. However, in places with intermediate environmental conditions, there are intermediate forms, and the two species are essentially identical except for a few characters that may be expected to vary in response to environmental factors. Subsp. chihuahuana tends to be shorter, to grow more towards the country's interior, where sites are drier and climate is more continental, has somewhat stiffer needles, 3 per fascicle, and somewhat larger cones, and from a distance the needles have a sort of "brushed" appearance, as if the tree was in a painting. P. leiophylla has 5 needles per fascicle, doesn't have the brushed appearance, and tends to occur as bigger trees, closer to the ocean, in stands with higher basal area. These are trees of the mountains, which means that the transition from maritime to continental climate happens very quickly, within a few kilometers of the crest of the range. Thus there is a relatively small area within which intermediate-form trees can be found. Both subspecies have early-deciduous fascicle sheaths, and are the only Mexican hard pines to have this character. Both subspecies also have a three-year cone maturation, unique in Latin American pines (but also seen in Pinus pinea); the ability to coppice (resprout from a cut stump), shared with Pinus rigida; and the common production of epicormic shoots, shared with a variety of species (photo on the Pinus canariensis page) (pers. obs. 2007, Perry 1991, and Farjon and Styles 1997).
Citations
Cruz-Nicolás, Jorge, Juan Pablo Jaramillo-Correa, and David S. Gernandt. 2024. Stochastic processes and changes in evolutionary rate are associated with diversification in a lineage of tropical hard pines (Pinus). Molecular Phylogenetics and Evolution 192:108011, https://doi.org/10.1016/j.ympev.2024.108011.
Gernandt, David S., Xitlali Aguirre Dugua, Alejandra Vázquez-Lobo, Ann Willyard, Alejandra Moreno Letelier, Jorge A. Pérez de la Rosa, Daniel Piñero, and Aaron Liston. 2018. Multi-Locus phylogenetics, lineage sorting, and reticulation in Pinus subsection Australes. American Journal of Botany 105(4):711–25. https://doi.org/10.1002/ajb2.1052.
López-Upton, Javier. 2003. Pinus leiophylla. Species description in the Tropical Tree Seed Manual. Available http://www.rngr.net/Publications/ttsm/Folder.2003-07-11.4726 (accessed 2007.08.31).
Rodríguez‐Banderas, Abril, Carlos F. Vargas‐Mendoza, Anna Buonamici, and Giovanni G. Vendramin. 2009. Genetic diversity and phylogeographic analysis of Pinus leiophylla: a post‐glacial range expansion. Journal of Biogeography 36(9):1807-1820.
Schlechtendal, D. F. L. von, and A. von Chamisso. 1831. Plantarum mexicanarum a cel. viris Schiede et Deppe collectarum (recensio brevis). Addenda (1). Linnaea 6:352-384 (p. 354).
See also