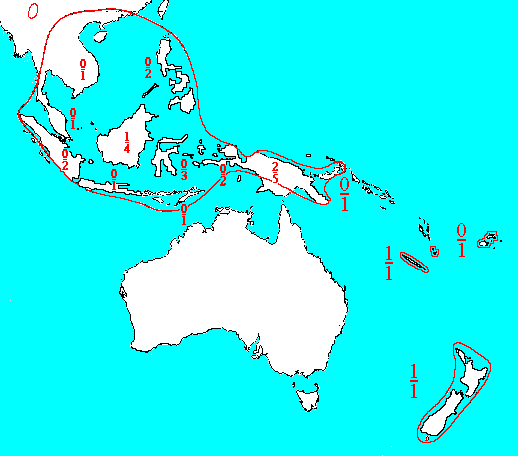
Range of the genus Dacrycarpus. Figures above the hyphen indicate the number of endemic species, those below the hyphen the total number of species. Redrawn from de Laubenfels (1988).
![]()
Range of the genus Dacrycarpus. Figures above the hyphen indicate the number of endemic species, those below the hyphen the total number of species. Redrawn from de Laubenfels (1988).
![]()
Dacrycarpus
Syn: Podocarpus sect. Dacrydioideae Bennett 1838; Podocarpus sect. Dacrycarpus Endl. 1847; Podocarpus sect. Dacrydium Bertrand 1874. Type species Dacrycarpus imbricatus (Blume) de Laubenfels 1969; basionym Podocarpus imbricatus Blume 1827" (de Laubenfels 1988).
This genus contains nine species, most of which were segregated from Podocarpus when the genus was described by de Laubenfels (1969):
"Dioecious shrubs or trees, to 41 m tall. Bark hard, dark brown or blackish but weathering to gray, surface rough with occasional lenticels, inside pink to reddish brown and granular or slightly fibrous, on older trees breaking off in small thick, somewhat vertically elongated plates or sometimes short strips. Leaves amphistomatic, spirally placed, broadly decurrent, apiculate. Leaves on primary shoots as well as on the basis of foliage shoots and fertile structures bifacially flattened, keeled on the dorsal side, lanceolate or sometimes triangular, often nearly appressed, mostly 1.5-3 by 0.5-0.8 mm, but wider on the decurrent part, up to at least 4 mm long on young plants and mostly 1-2 mm long at the base of foliage shoots or fertile structures. The apex of resting shoots a loose cluster of reduced leaves. Juvenile type of leaves on special shoots that generally do not continue growth after reaching a resting stage, bilaterally flattened and usually slightly keeled on both faces, falcate and then curved forward towards the apex so that the apiculate tip is oriented more or less parallel with the shoot, otherwise linear or less often lanceolate, spreading at about a 60 angle, the centre of the leaf either straight or gradually curving to the tip, distinctly shorter towards either end of the shoot so that the whole shoot has a feather-like appearance, mostly 6-12 mm long and c. 1 mm wide, usually distichous, gradually changing to an adult form but often fertile at intermediate stages of this transition. Final adult forms not distichous, generally shorter and more robust than the juvenile leaves, more or less uniform along the shoot, the apiculate tip still bent forward parallel to the shoot, in some species nearly identical with the leaves of primary shoots. Fertile structures terminal on short, mostly lateral shoots, the seed-bearing structures usually on a considerably longer shoot than that of the pollen cone. Immature pollen cones at first sometimes nearly spherical, then becoming somewhat elongated but finally elongating abruptly with a slight decrease in diameter at maturity, then mostly c. 6-10 mm long and 2-3 mm diam., sometimes longer. Apex of microsporophyll triangular, acute to apiculate, c. 1.2 by 0.8 mm. Shoots for female structures 3-17 mm long. Leaves at the base of the seed-bearing structure sharply elongated to form an involucre which often surrounds the immature seed-bearing structure but which in the shorter examples becomes spreading as the structure grows. Seed-bearing structure composed of a small warty receptacle, 2.5-4 mm long, 2.5 mm diam., which becomes greatly enlarged, fleshy, and first orange then red or in other species purple when ripe, later turning brown, bearing one or two protruding short sterile leaf-like bracts and one or two subterminal fertile bracts. The inverted ovules completely surrounded by the epimatium and fused as a rib along one side with the fertile bract whose short free tip forms a small off-centre crest over the mature structure. Mature seed nearly spherical (or oval) but remaining covered by the leathery epimatium and scale, forming an erect or somewhat oblique structure which in most species is c. 5-6 mm long and 4.5-5.5 mm diam., dark in colour" (de Laubenfels 1988).
"Note. Sterile specimens strongly resemble Dacrydium and hence the generic name. The fusion of the fertile scale with the epimatium is a unique trait of the genus while the seeds of Dacrydium are furthermore naked. In most cases sterile specimens can be readily distinguished from Dacrydium by the distinctly dimorphic foliage" (de Laubenfels 1988).
N Burma and S China to Fiji and New Zealand, with greatest diversity in New Guinea (5 spp.) (de Laubenfels 1988).
Dacrycarpus dacrydioides is both largest and tallest. It is also the only species for which I have found any age data; approximately 716 years.
Data are scanty; see the species descriptions.
See the species descriptions.
The genus name is derived from the Greek words for "tears" and "fruit", in reference to the shape of the seed cones.
"Fossils are known from N. Antarctica (Graham Land) and New Zealand (Middle Jurassic) and S. Patagonia (Upper Cretaceous to Oligocene) where the genus is now extinct. In the Eocene found in New Zealand, and since the Oligocene also in SE. Australia, where it became extinct (Florin, Kongl. Svensk. vet. Ak. Handl. 111, 19, n. 2, 1940, 70; Acta Horti Berg. 20 (4), 1963, 188, f. 19: map)" (de Laubenfels 1988).
Laubenfels, D. J. de. 1969. A revision of the Malesian and Pacific rainforest conifers, I. Podocarpaceae, in part. Journal of the Arnold Arboretum 50:315-317. Available: Biodiversity Heritage Library, accessed 2021.12.24.
Last Modified 2023-02-26