Araucaria
Juss. 1789
Common names
Araucaria.
Taxonomic notes
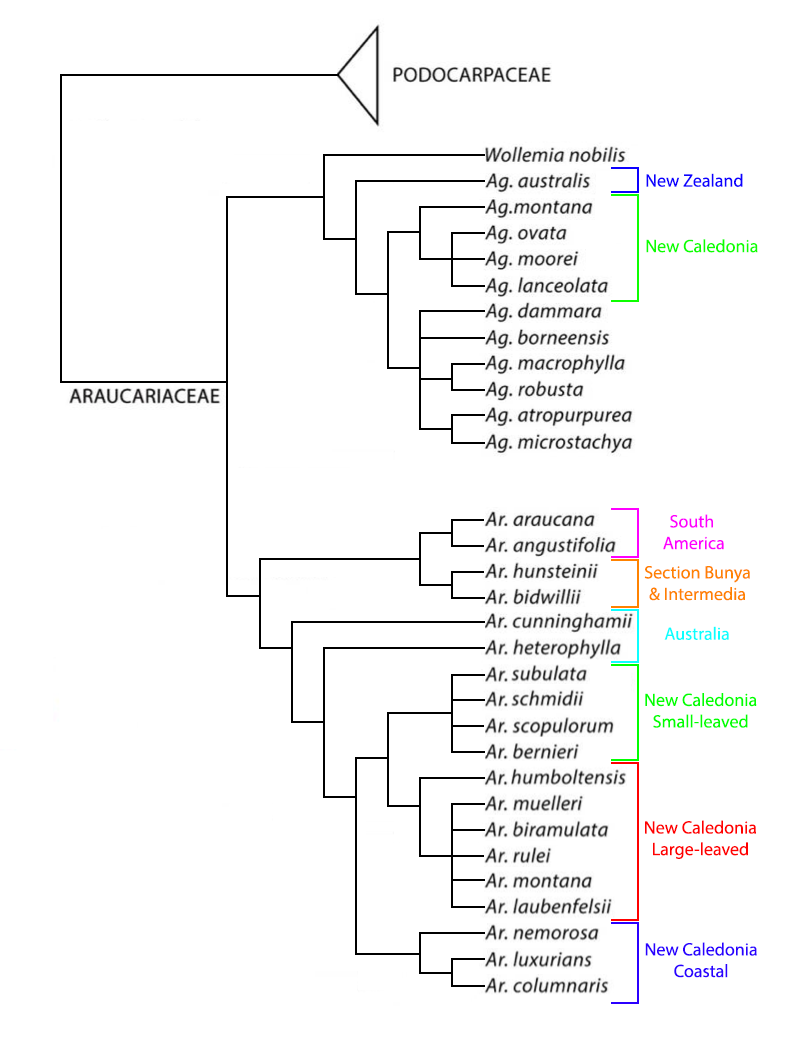
In this treatment there are 20 species. The extant species of the genus are assigned to 3 sections as follows (Wilde and Eames 1952, except section Intermedia is reduced to synonymy with section Bunya in accordance with the molecular analyses of Escapa and Catalano [2013]):
| | |
Sect. Araucaria (syn: Colymbea): |
A. araucana and A. angustifolia, both native to South America. |
Sect. Bunya: |
A. bidwillii and A. hunsteinii. |
Sect. Eutacta: |
A. cunninghamii, A. heterophylla, and the 14 New Caledonia endemics: A. bernieri, A. biramulata, A. columnaris, A. goroensis, A. humboldtensis, A. laubenfelsii, A. luxurians, A. montana, A. muelleri, A. nemorosa, A. rulei, A. schmidii, A. scopulorum and A. subulata. |
| | |
Besides these, Ohsawa et al. (1995) have also described one extinct section, Yezonia, containing one extinct species (A. vulgaris) from the Cretaceous of northern Japan (many extinct species have been described in the other sections as well). Although the sections were defined largely on the basis of morphological data, a series of more detailed analyses including multiple sources of molecular data, fossil evidence, and morphological characters was used to develop the cladogram shown here, redrawn from an original by Escapa and Catalano (2013) and using information from the molecular analysis of New Caledonian species by Gadeul et al. (2012). Analysis of the rbcL gene from chlorplast DNA of the species of Section Eutacta, endemic to New Caledonia, revealed nearly identical rbcL sequences, suggesting relatively recent and rapid adaptive radiation, probably driven by the island's extraordinary edaphic conditions, i.e., ultramafic soils (Setoguchi et al. 1998). The analysis by Gadeul et al. (2012) used 142 amplified fragment length polymorphism (AFLP) markers from 71 samples to evaluate all of the New Caledonian species in an analysis that found strong support for three major groups: a coastal group including A. columnaris, A. luxurians and A. nemorosa; a group of small-leaved montane species including A. bernieri, A. schmidii, A. scopulorum and A. subulata; and a group of large-leaved montane species including the remaining taxa, likely including A. goroensis, which had not been described at the time of the study. The three clades did not show a clear environmental association and reflect speciation from differential adaptation and vicariance; the results also pointed towards possible presence of cryptic species, a prediction confirmed a few years later when several of the authors described A. goroensis.
Syn.: Eutassa Salisb. (applied only to the species of Section Eutacta).
Description
Evergreen trees with regularly whorled branches. Buds inconspicuous. Young trees branched to the ground, old trees with a long clear bole and flattened crown. Juvenile leaves needle-like, thin, loosely imbricate, spirally arranged. Adult leaves usually scale-like or sometimes needle-like, spirally arranged or nearly 2-ranked, usually closely imbricate, overlapping, flattened, sometimes lanceolate and sharp-pointed, persistent for many years, often with faint stomata above. Male cones terminal and solitary or disposed in clusters, with numerous spirally arranged stamens, anthers 6-8 celled. Female cones globose, ripening in 2-3 years, heavy, milky; scales usually with thin spiny apical umbos, scales thin and somewhat flattened, cones dehiscent at maturity; ovules one per scale. Ovules and seeds united with the scale. Seeds usually winged on the edges, sometimes edible. Cotyledons 2-4. Germination with epigeal or hypogeal cotyledons (Silba 1986).
Distribution and Ecology
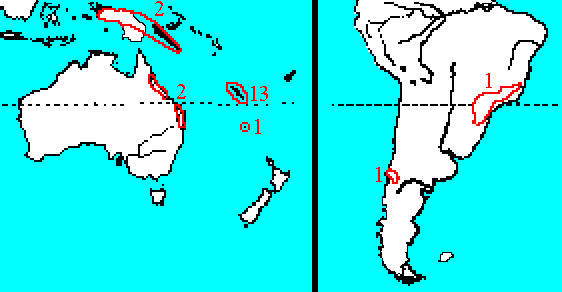
Araucaria has a highly disjunct distribution: Chile, Argentina, southern Brazil, New Caledonia, Norfolk Island, Australia, and New Guinea. Some authors regard its occurrence in South America as the result of a vicariance event, but considering the ancient lineage of the family and its former abundance in the Northern Hemisphere, its extant distribution is likely relictual (Setoguchi et al. 1998).
This map shows herbarium records of Araucaria species native to New Caledonia. Letter corresponds to first letter of species epithet (A. goroensis coded as A. muelleri, see A. goroensis for discussion) with different colors for species having same first letter. Click on an icon for further information. Distribution data from GBIF (2020.03.30), edited to remove duplicates. See Araucariaceae for a map showing the global distribution of Araucaria.
Remarkable Specimens
Araucaria araucana grows to the largest size. A. bidwillii grows nearly as large. Few data are available for most species, but none of the New Caledonian species appear to grow exceptionally tall or thick. The tallest is almost certainly A. hunsteinii, reported to 89 m, almost 35 m taller than any other species (this height is not corroborated and is somewhat unbelievable).
As for the oldest species, I have heard that A. araucana may exceed 1000 years. There are no data for most species, but A. araucana is also distinctive within the genus in occupying relatively dry habitats, which are typical for most extremely old conifers.
Ethnobotany
Many species, due to their large size, are or have been important sources of timber. Examples include A. angustifolia, A. araucana, A. bidwillii, A. cunninghamii and A. hunsteinii. The nutritious and large seeds of A. bidwillii and A. angustifolia were an important food source for aboriginal peoples and remain a popular delicacy.
There have been some dendrochronological studies with A. araucana and A. nemorosa.
Observations
Some of the more impressive (large) species are popular ornamentals, widely planted in warm temperate and subtropical regions. Quite a few species may be seen in many warm temperate or tropical botanical gardens. Species native to temperate and subtropical regions in Chile, Argentina, Brazil, and Australia have been preserved in some public parks or forest preserves, where they may be found in their natural habitat. However, many of the tropical species, such as the 14 species endemic to New Caledonia and the Malesian species, have received limited protection, are difficult to locate in the wild, and are threatened with extinction. There is a trend toward recognizing the botanical significance of these tropical species and providing them with statutory protection, but at the same time their habitat is rapidly disappearing due to population and development pressures. Their fate is for the most part uncertain.
Remarks
The genus is named after the province of Arauco in South Chile (Vidakovic 1991).
From Setoguchi et al. (1998): "New Caledonia, one of the continental islands, originated from a part of Gondwanaland and was separated from the continent between the Triassic and the end of the Jurassic. However, a large area of New Caledonia is covered with ultramafic soil derived from peridotite, and it is suggested that this area was formed at the end of the Eocene. Most of the species of Araucariaceae are distributed in this soil (12 of the 13 species in Araucaria and four of the five species in Agathis). Jaffre suggested that there was differentiation of new species of Araucaria in the post-Eocene, after the emplacement of peridotite. His discussion agrees with the findings of very low sequence divergence in the rbcL gene among endemics of Araucaria and Agathis. It is suggested that the adaptive radiation to new edaphic conditions, i.e., ultramafic soils, caused their rapid differentiation after the Eocene."
Abstract of Haines et al. (1984): "Pollination was studied in three species of Araucaria: A. cunninghamii, A. heterophylla and A. bidwillii. More limited observations were made for A. hunsteinii and A. angustifolia. The investigation revealed considerable variation among species of Araucaria with respect to pollination features. The bract scale of the receptive cone of both A. cunninghamii and A. heterophylla has a stoma-free furrow which directs pollen grains onto a thin flange on the tip of the ovuliferous scale. The bract scale of A. bidwillii bears stomata over most of its surface, and is broady scalloped so that pollen grains are deposited in a band across both bract and ovuliferous scales. On germination, the pollen tubes of these three species penetrate the epidermis of the scale and for a short distance grow beneath the surface before emerging and travelling, without branching or major deviation, towards the proximal end of the ovuliferous scale. Results are discussed in relation to pollination mechanisms in other conifers and to taxonomic divisions within the Araucariaceae."
Citations
Escapa, I. H., and S. A. Catalano. 2013. Phylogenetic analysis of Araucariaceae: Integrating molecules, morphology, and fossils. International Journal of Plant Sciences 174(8):1153–1170.
Gaudeul, M., G. Rouhan, M. F. Gardner, and P. M. Hollingsworth. 2012. AFLP markers provide insights into the evolutionary relationships and diversification of New Caledonian Araucaria species (Araucariaceae). American Journal of Botany 99(1):68–81.
Haines, R.J.; Prakash, N. and Nikles, D.G. 1984. Pollination in Araucaria Juss. Australian Journal of Botany 32(6): 583-594.
Hortus Botanicus Catinensis, URL = http://www.dipbot.unict.it/orto/0001-1.html.
Jussieu, A. L. de. 1789. Genera Plantarum, p. 413. Available: Biodiversity Heritage Library, accessed 2023.03.04.
Ohsawa, T., H. Nishida and M. Nishida. 1995. Yezonia, a new section of Araucaria (Araucariaceae) based on permineralized vegetative and reproductive organs of A. vulgaris comb. nov. from the upper Cretaceous of Hokkaido, Japan. Journal of Plant Research 108: 25-39.
[Anonymous, undated]. Pinheiro-do-Paraná, http://www.pr.gov.br/seec/pinheiro/index.html (accessed 2000.06.23, now defunct).
Schmid 1981.
Wilde, M.H. and A. J. Eames. 1952. The ovule and seed of Araucaria bidwilli with discussion of the taxonomy of the genus. II. Taxonomy. Annals of Botany New Series 16: 27-47.
See also
Bose, M.N. 1975. Araucaria haastii Ettingshausen from Shag Point, New Zealand. Palaeobotanist 22: 76-80.
Escapa, I. H. and S. A. Catalano. 2013. Phylogenetic analysis of Araucariaceae: integrating molecules, morphology and fossils. Int. J. Plant Sci. 174(8):1153-1170.
Stockey, R.A. 1975. Seeds and embryos of Araucaria mirabilis. American Journal of Botany 62: 856-868.
Stockey, R.A. 1978. Reproductive biology of Cerro Cuadrado fossil conifers: Ontogeny and reproductive strategies in Araucaria mirabilis (Spegazzini) Windhausen. Palaeontographica B 166: 1-15.
Stockey, R.A. 1980a. Anatomy and morphology of Araucaria sphaerocarpa Carruthers from the Jurassic inferior Oolite of Bruton, Somerset. Botanical Gazette 141: 116-124.
Stockey, R.A. and H. Ko. 1986. Cuticle micromorphology of Araucaria de Jussieu. Botanical Gazette 147: 508-548.
Stockey, R.A., M. Nishida, and H. Nishida. 1992. Upper Cretaceous araucarian cones from Hokkaido: Araucaria nihongii sp. nov. Review of Palaeobotany and Palynology 72: 27-40.
Stockey, R.A., M. Nishida, and H. Nishida. 1994. Upper Cretaceous araucarian cones from Hokkaido and Saghalien: Araucaria nipponensis sp. nov. International Journal of Plant Science 155: 800-809.
Veillon, J.-M. 1978. Architecture of the New Caledonian species of Araucaria. P. 233-245 in P.B. Tomlinson and M.H. Zimmermann [eds.], Tropical trees as living systems. Cambridge University Press, Cambridge.
Veillon, J.-M. 1980. Architecture des especes neo-caledoniennes du genre Araucaria. Candollea 35: 609-640.