Gymnosperms
[colloquial, first use ca. 1703 by John Ray]
Common names
Gymnosperms (from the Greek, γυμνόσπερμος, meaning "naked seed" because the seeds do not develop within an ovary).
Taxonomic notes
The following table describes the relative hierarchy of gymnosperm taxonomy and provides links to descriptions of each order, family, and genus.
| Order |
Family |
Genera |
Species |
| Cycadales |
Cycadaceae |
Cycas |
120 |
| Zamiaceae (incl. Stangeriaceae) |
Bowenia, Ceratozamia, Chigua, Dioon, Encephalartos, Lepidozamia, Macrozamia, Microcycas, Stangeria, and Zamia |
246 |
| Ginkgoales |
Ginkgoaceae |
Ginkgo |
1 |
| Gnetales |
Welwitschiaceae |
Welwitschia |
1 |
| Gnetaceae |
Gnetum |
40 |
| Ephedraceae |
Ephedra |
71 |
| Pinales |
Pinaceae |
Abies, Cathaya, Cedrus, Keteleeria, Larix, Nothotsuga, Picea, Pinus, Pseudolarix, Pseudotsuga, and Tsuga |
234 |
| Araucariales |
Araucariaceae |
Agathis, Araucaria, and Wollemia |
41 |
| Podocarpaceae (incl. Phyllocladaceae) |
Acmopyle, Afrocarpus, Dacrycarpus, Dacrydium, Falcatifolium, Halocarpus, Lagarostrobos, Lepidothamnus, Manoao, Microcachrys, Nageia, Parasitaxus, Pectinopitys, Pherosphaera, Phyllocladus, Podocarpus, Prumnopitys, Retrophyllum, Saxegothaea, and Sundacarpus. |
174 |
| Cupressales |
Sciadopityaceae |
Sciadopitys |
1 |
| Cupressaceae (incl. Taxodiaceae) |
Athrotaxis, Austrocedrus, Callitris, Callitropsis, Calocedrus, Chamaecyparis, Cryptomeria, Cunninghamia, Cupressus, Diselma, Fitzroya, Glyptostrobus, Hesperocyparis, Juniperus, Libocedrus, Metasequoia, Microbiota, Papuacedrus, Platycladus, Sequoia, Sequoiadendron, Taiwania, Taxodium, Tetraclinis, Thuja, Thujopsis, Widdringtonia, and Xanthocyparis |
154 |
| Taxaceae (incl. Cephalotaxaceae) |
Amentotaxus, Austrotaxus, Cephalotaxus, Pseudotaxus, Taxus, and Torreya |
28 |
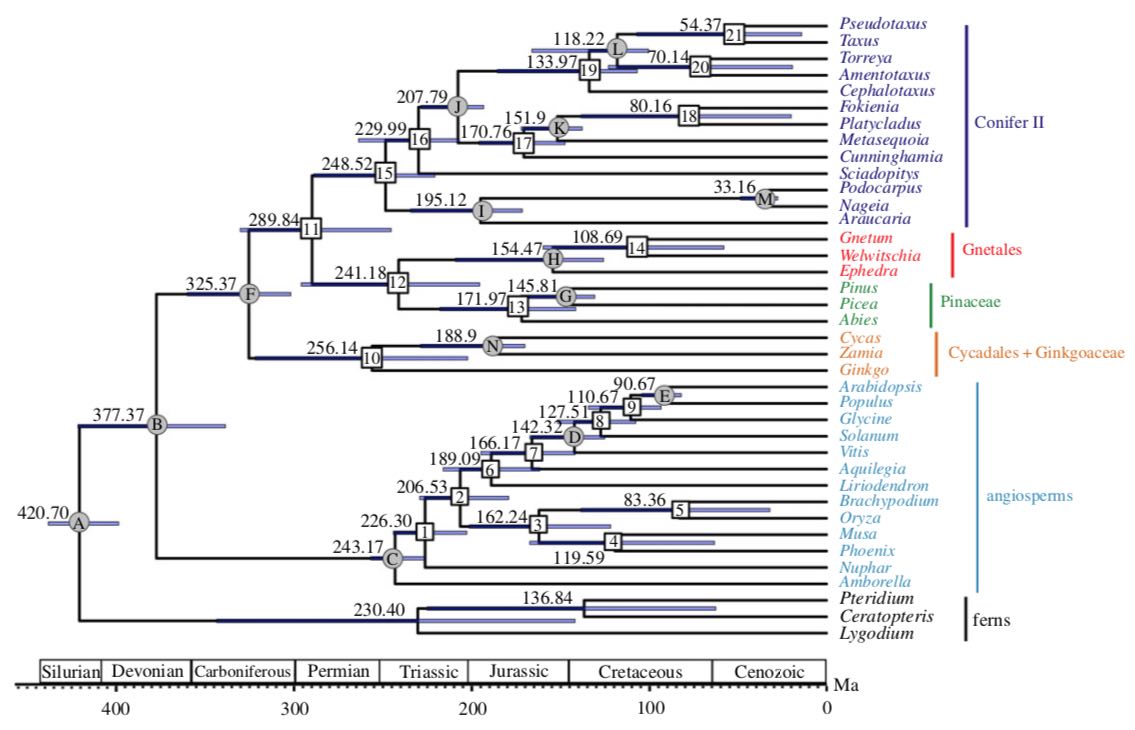
This database provides information on taxa belonging to the gymnosperms. The gymnosperms are a relatively small and highly distinctive assemblage of plants that are for the most part very ancient in lineage, but which have great cultural and ecological importance at a global scale—a significance far greater than their species diversity alone would imply. The gymnosperms have long been recognized as a distinctive group of plants, but for a VERY long time (more than a century) there was active debate about whether the principal groups within the gymnosperms share a common ancestor, and if so, whether that common ancestor was shared with the flowering plants, also called angiosperms. A succinct online review of the subject is provided by Castillo (2001), and an excellent further review by Wang and Ran (2014). This latter review is one of the best survey articles about gymnosperms that I have found. In 2018, gene sequencing technology had finally developed to the point where it was possible to confidently determine which parts of the plant genome were fundamental to answering this question, and to fully sequence representatives of a large number of relevant species. This work, reported by Ran et al. (2018), strongly supports the long-standing "gnepine" hypothesis; the resulting cladogram, shown at right, divides the traditional conifers into two clades, one containing the Pinaceae and the other all remaining conifer families.
The Pinaceae are sister to the gnetophytes (Ephedra, Gnetum and Welwitschia), in a clade sister to the remaining conifers, and these collectively are sister to Ginkgo and the cycads, all of which together are sister to the angiosperms. As best I can determine, the terminology has not yet caught up with this idea, so there is not yet a term for the non-Pinaceae conifers, or for the Gnetales+Pinales clade; the table above will have to serve for now (2021). This also leaves us with the peculiar conclusion that the Gymnosperms are a monophyletic group, but the conifers are not! Some authorities remain sceptical of the gnepine hypothesis, noting the general lack of morphological support for it. However, morphology has limited utility to explain relationships between orders or classes among any group of plants, so I don't find this objection problematic. A more serious criticism is that "Because of extinction, molecular datasets poorly sample diversity from periods much earlier than the Late Cretaceous. This fundamentally limits their utility for understanding deep patterns of character evolution and resolving the overall pattern of conifer phylogeny" (Leslie et al. 2018). This caveat shows that we might always have limited confidence about the deep structure of gymnosperm phylogeny.
As of 2022, full genome sequencing has been performed for a representative of each major group of gymnosperms. These studies have provided considerable insight about deep structure within each group. The gnetales have the smallest genomes at about 4 Gb, the cycads are about 8 Gb, ginkgo is near 12 Gb, and both the pines and the other conifers have genomes of about 20 Gb (range: 6 to 37 Gb). These genomes code for about 20,000 to 50,000 genes, and typically contain a large fraction of repetitive DNA. This high fraction of repetitive DNA may explain the relatively large genome size, compared to most angiosperms. Although the gymnosperms have traditionally been thought of as a primarily diploid group (with some polyploidy at the species level), evidence of ancient whole-genome-duplication events has been found each major gymnosperm group (cycads, non-Pinaceae conifers, Pinaceae, and gnetophytes); these events have likely been associated with episodes of rapid evolutionary change (Ahuja and Neale 2005, Li et al. 2015, Liu et al. 2022).
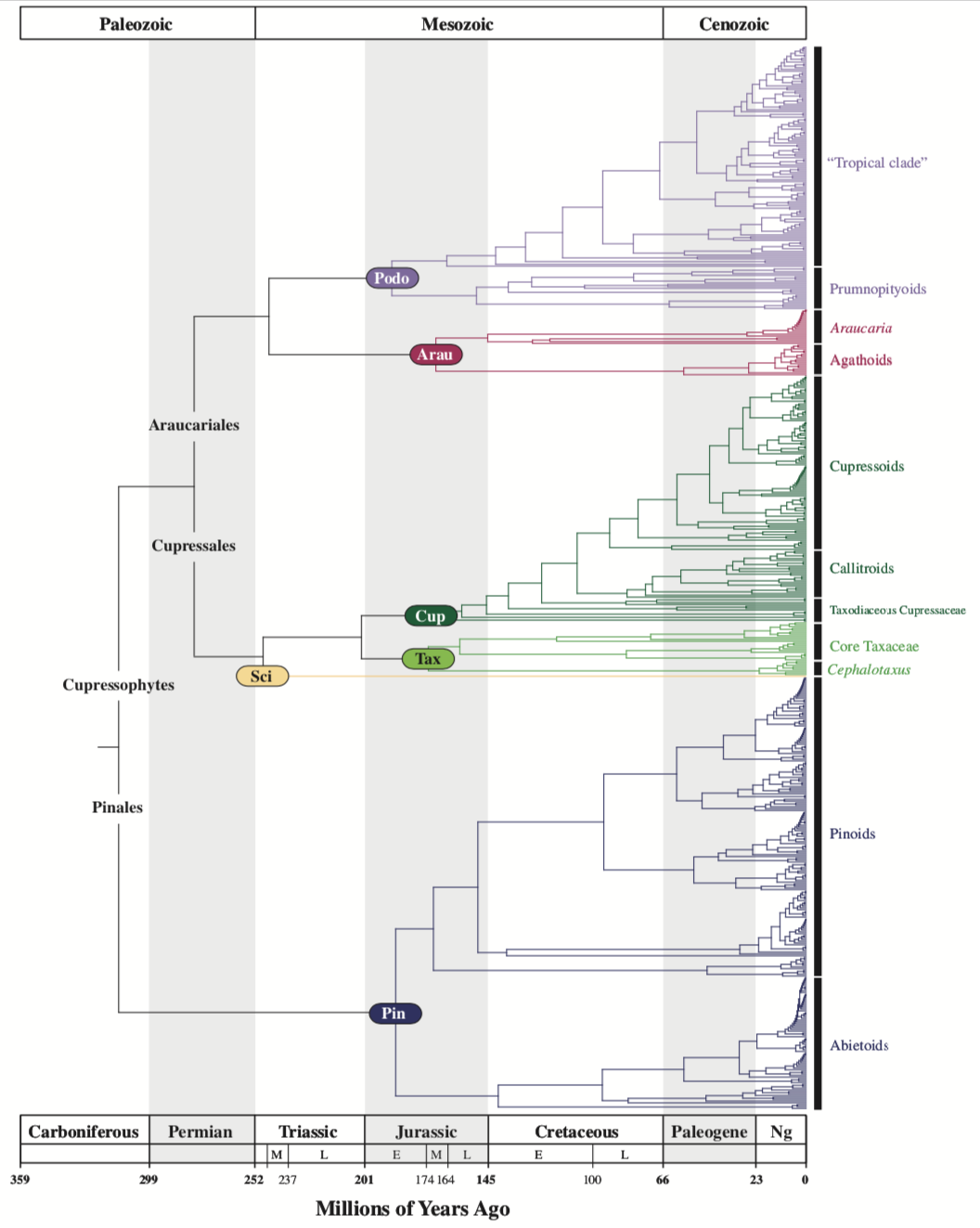
Although molecular studies have done much to improve our understanding of evolutionary relationships between extant gymnosperms, changing relationships at evolutionary timescales are best elucidated through the fossil record. The following points summarize the chronology of gymnosperm evolution, as does the figure at right from Leslie et al. (2018):
- Late Devonian (385-359 million years ago [my]): Earliest seed plants arise (Hill 2005).
- Middle Pennsylvanian (310 my): Earliest conifers, cycads, and Ginkgo arose at about the same time, perhaps from a precursor in the Cordaitales, a plant that bore seeds in conelike structures. These earliest gymnosperms were not referrable to any families now existing. The Cordaitales persisted until the mass extinction at the end of the Triassic (201 my) (Hilton 2014).
- Permian (299-251 my): First appearance of Ginkgoales. The group is most diverse and abundant during the Jurassic (Thomas and Spicer 1986 in Hill 2005).
- Lower Permian (299-271 my): More recognizable cycads appear, e.g. Crossozamia (Gao and Thomas 1989); cycad diversity and abundance peaked during the Mesozoic (Hill 2005).
- Middle Permian (271-260 my): First evidence of Gnetophytes, limited to pollen evidence (Crane 1988); macrofossils attributable to Gnetophytes appear only much later (Cretaceous).
- Upper Triassic (235-202 my): First appearance of the Bennettiales. This was one of the most abundant and diverse gymnosperm groups in the Mesozoic, although it became extinct (along with the dinosaurs) at the end of the Cretaceous. Evolutionary relationships are unclear but the Bennettiales may have been most closely related to the Gnetales (Hill 2005).
- Triassic, earliest Norian (228-225 my): The first appearance of Podocarpaceae, the extinct species Rissikia media (Townrow 1967 in Rothwell et al. 2012).
- Lower Jurassic (202-176 my): First appearance of Ginkgo, the oldest extant conifer genus (Tralau 1968)
- Jurassic, Sinemurian (197-190 my): First record of Cupressaceae, the extinct species Austrohamia minuta (Escapa et al. 2008 in Rothwell et al. 2012).
- Jurassic, Pliensbachian (190-183 my): First record of Taxaceae, the extinct species Paleotaxus rediviva (Nathorst 1908 in Rothwell et al. 2012).
- Jurassic, Pliensbachian (190-183 my): First record of Araucariaceae, the extant genus Araucaria (Arrondo and Petriella 1980 in Rothwell et al. 2012).
- Jurassic, Kimmeridgian (156-151 my): First record of Pinaceae, the extinct species Eathiestrobus mackenziei (Rothwell et al. 2012).
- Lower Cretaceous (130 my): First appearance of Pinus, the extinct species Pinus yorkshirensis from a fossil cone preserved in the Wealden Formation in Yorkshire, UK (Ryberg et al. 2012).
- Cretaceous, Aptian (125-112 my): First Gnetales macrofossils, in the form of Drewria potomacensis, an extinct species with leaves resembling those of Gnetum (Crane and Upchurch 1987 in Hill 2005).
- Cretaceous, Conacian (89-86 my): First record of Sciadopityaceae, the extant genus Sciadopitys (Ohsawa et al. 1991 in Rothwell et al. 2012).
Although morphologically, ecologically and taxonomically diverse, and individually very numerous, the gymnosperms are not represented by a great many species. In this treatment there are 1107 species (as of 2021.12.10), or about as many as may be found in the largest genera of flowering plants (such as Acacia). Of this total, one species represents the ginkgo, 112 represent the gnetophytes, 366 are cycads, and the remaining 628 species are conifers. Since I started tracking these numbers in about 1985, the numbers of gnetophytes and cycads have increased substantially (there used to be about 60 gnetophytes and 200 cycads), while the number of conifers has held fairly constant.
I am sometimes asked about the distinctions between genera, species, subspecies and varieties. Among the gnetophytes and cycads, I largely accept the "normal" taxonomy, which is currently defined for most of the world's botanists by the Plants of the World Online database. Among conifers, I try to track the literature closely and critically evaluate new information, which in the 21st century has mostly come from gene sequencing studies. These are referenced in the "Taxonomic notes" for each taxon. In general, morphologically distinct genera in the conifers are marked by conspicuous differences in the morphology of the seed cone. Distinct species are generally marked by conspicuous differences in foliage characters (and usually by other points of morphology as well), and by different geographic and ecological affiliations that provide for reproductive isolation and niche specialization. Distinct subspecies are generally also marked by different geographic and ecological affiliations, but morphological characters may be quite subtle, and reproductive isolation may be weak or absent. Distinct varieties are generally marked by observable morphological differences that may or may not have ecological consequences. Application of these definitions results in most infraspecific taxa being described as subspecies.
Description
Plants are classified into two large groups, vascular plants and nonvascular plants. Nonvascular plants lack tissues that have been specialized for the transport of water and nutrients between distant parts of the plant. They include all algae, mosses, and their allies. Vascular plants have such transport systems. They include the vast majority of familiar terrestrial plants and can be divided into four major groups according to major features of their reproductive systems. The simplest and oldest of these is Lycopodiopsida or lycopods (about 1,300 species), which consist mostly of the club mosses. Slightly later, but still a largely relict group, are the Polypodiopsida or ferns and fern allies (about 11,000 species). The next group is the Pinophyta or Gymnospermae (about 1,100 species), which are the focus of this presentation. The last and by far the largest group (about 300,000 species), is the Angiospermae or flowering plants.
Gymnosperms are woody plants, either shrubs, trees, or, rarely, vines (some gnetophytes). They differ from flowering plants in that the seeds are not enclosed in an ovary but are exposed within any of a variety of structures, the most familiar being cones. The gymnosperms display four architectural models of tree growth, which determine the basic form and appearance of the plant; see Tomlinson (1983) for more detail. A more anatomically detailed discussion of the comparative morphology of gymnosperms and related vascular plants can be found in the On-Line Biology Book or by clicking on Google Search (Gymnosperm Life Cycle). More detailed descriptions are provided for each gymnosperm family.
Gymnosperms are variably monoecious or dioecious, and about 1% of species (mostly junipers) are either, or can switch between monoecious or dioecious. The dioecious species include all cycads, all gnetophytes, ginkgo, 95% of the Podocarpaceae, 93% of the Taxaceae, and about 30% of the Cupressaceae (all are junipers). In general these plants live in warm climates and put considerable energy into producing fairly large seeds or seed-bearing structures that are attractive to animals, which are the primary dispersers of those seeds. Overall 65% of gymnopsperms are dioecious, a dramatic contrast to 5% of angiosperms. The monoecious species, on the other hand, include almost all Araucariaceae and Pinaceae, and 70% of the Cupressaceae; they are predominately wind-pollinated with wind-dispersed seeds, and live in temperate to cold climates (Walas et al. 2018).
It seems odd, but apart from the naked seed, the gymnosperms really do not share many attributes. Wood and leaf, habit and habitat, physiology and anatomy - all are quite variable between the four major groups. We might have chosen to put "the gymnosperms" together in one group because they are all relicts, survivors of an ancient flora. They all seem strange to children of the Cenozoic savannas.
Distribution and Ecology
Living gymnosperms are distributed worldwide, excepting polar regions, the highest mountains, the driest deserts, and a few oceanic islands. The cycads are primarily tropical and subtropical, with greatest diversity in Central America, Africa and Australia. Ginkgo is confined to a very small native range in China. Welwitschia occurs only in the subtropical desert of coastal Namibia, while the Ephedraceae are very widespread in generally semiarid, subtropical to cold-temperate habitats in Africa, Asia, Europe and the Americas. Gnetum is found in the wet tropics; in Indomalaysia, tropical parts of West Africa, Fiji, and the northern regions of South America.
The conifers are cosmopolitan. Species in the Pinaceae are almost entirely confined to the northern hemisphere, in habitats ranging from wet tropics to boreal forests to very dry, cold mountains. The Podocarpaceae and Araucariaceae are most widespread in the southern hemisphere and tropical northern latitudes, generally occupying warm and mesic habitats. The Taxaceae are very widespread, although nowhere abundant, in the northern hemisphere and a bit of the southern, generally in warm to cool mesic habitats, though a few species occur in the wet tropics. The Sciadopityaceae is only native in Japan, in a mesic temperate climate. The Cupressaceae have the broadest distribution and the widest climate tolerances of any gymnosperm family; they are abundant in Africa, the Americas, Australia, Europe, and Asia, and occasional on oceanic islands. They occupy habitats from dry deserts to temperate rainforests, in climates ranging from wet-tropical to boreal-continental.
Worldwide distribution of conifers, generalized to 1x1 degree cells; data from the BRAHMS database at Conifers of the World, accessed 2021.11.11. Currently no comparable mapping has been prepared for other gymnosperms.
Gymnosperms, generally speaking, are of serious conservation concern and have long been recognized as such. A recent IUCN (2020) summary found that a large fraction of gymnosperm taxa, including a third of all conifers and nearly two-thirds of all cycads, are classified as of conservation concern:
| Group |
Critically Endangered |
Endangered |
Vulnerable |
Subtotal |
Percentage of group at risk |
| Conifers |
29 |
96 |
80 |
205 |
34% |
| Cycads |
53 |
65 |
74 |
192 |
63% |
| Gnetophytes |
0 |
1 |
3 |
4 |
4% |
Besides the species inventoried in this table, the monotypic taxa Ginkgo biloba and Welwitschia mirabilis are also generally regarded as endangered. The reasons for this situation are variable, but in all major groups the loss of habitat through conversion to developed cover types (agriculture, pasture, urbanization) is the leading factor of decline. In many species, contributing factors include harvest, introduced pests and diseases, and climate change. Cycads, especially, are also harmed through illegal removal by collectors.
Although only a minority of species have been investigated, nearly all gymnosperms appear to form mycorrhizas. Vesicular-arbuscular mycorrhizas are seen in ginkgo (Fontana 2006), cycads (Fisher and Vovides 2004), conifers excepting the Pinaceae (Brundrett 2008), and Ephedra (Collier et al. 2003). Ectomycorrhizas are seen in the Pinaceae (Brundrett 2008) and the Gnetaceae (Bechem and Alexander 2012, Tedersoo and Põlme 2012). Only Welwitschia is thought to be non-mycorrhizal (Jacobson 1993).
Remarkable Specimens
Large and Tall Trees
Ginkgo can grow very large, while the conifers include many of the largest trees on earth. Sequoiadendron giganteum (Cupressaceae) is the largest of all trees, but many other giants exist as well. A hundred years or so ago the largest was probably Sequoia sempervirens, also in the Cupressaceae, but the finest stands in that species were all destroyed by the loggers long ago; only remnants remain. Still, it is the second-largest conifer as measured by biomass, and the tallest of all, with many trees over 100 m tall. The third largest species is Agathis australis (Araucariaceae). Other giants include Pseudotsuga menziesii (Pinaceae), Podocarpus totara (Podocarpaceae), and Taxus sumatrana (Taxaceae). A summary listing of big trees is given in the sidebar for A Tale of Big Tree Hunting In California.
Old Trees
The age of a gymnosperm is usually difficult to determine. Precise numbers are only available if someone recorded the date of planting. Many conifers, and the ginkgo, have (approximately) annual rings, permitting approximate determination of the age of an individual, but in harsh environments many years may pass before the first ring is made. For example, Ken Lertzman (pers. comm. 1990) found mountain hemlock seedlings up to 18 years old (based on bud scar counts) that had yet to form their first ring. The greatest age known from ring counts is about 5000 years, for Pinus longaeva. It is often described as the oldest living thing. Several other species, such as Thuja plicata (Cupressaceae) and Taxus baccata (Taxaceae), might achieve comparable ages but we cannot tell because they live in moist climates where the tree's woody heart rots away while the rest of the tree is in the prime of life, so that no record of the rings is left behind. For further detail on the oldest known gymnosperms, see the article How Old Is That Tree?.
Many other trees, including most tropical species, and all non-tree gymnosperms, do not produce annual rings. Age estimates for such plants are usually based on extrapolation of observed growth rates. Such extrapolation may lead to estimation errors of several hundred percent (usually overestimation, because trees grow more slowly as they age).
Many species of plants are clonal, so that numerous different individuals are genetically identical. Among the oldest, a clone age of 43,600 years has been estimated for Lomatia tasmanica, a Tasmanian tree (DEWHA 2010). A clone age estimate of 10,500 years has been proposed for Huon pine, Lagarostrobos franklinii, and a comparable age may be reached in the redwood, Sequoia sempervirens, which is only one of many gymnosperms that can sprout from fallen stems or cut stumps. Some cycads produce clones by producing buds at the base, which may become separated from the main plant and then root. The oldest gymnosperm clones are likely to be found among species prone to vegetative reproduction in an area of extraordinarily stable climate, such as cycads of the genus Encephalartos growing in South Africa.
Dendrochronology, the study of tree rings, is necessarily restricted to trees that form annual rings. This includes hundreds of species; among gymnosperms, it includes Ginkgo and many, perhaps most conifers. I have not found dendrochronological research into Ginkgo, but nearly all conifers living in temperate or boreal climates have been investigated for use in dendrochronology, and most have proven suitable, with the exceptions found mainly in areas where extreme stress from cold, drought or other factors frequently prevents the formation of an annual ring, or among taxa where the ring is discontinuous around the circumference of the tree. Tropical gymnosperms have been less widely studied, but generally, those growing in climates with pronounced wet and dry seasons form annual rings and those with year-round rainfall are not usable.
Ethnobotany
Gymnosperm use by humans varies between gymnosperm groups:
- Cycads are a source of food; although many species are highly toxic, with proper preparation their seeds and/or stems are nutritious. Their leaves also provide a source of fiber and thatch.
- Ginkgo is a sacred tree, planted at many oriental temples, and it seeds particularly have a long history of medicinal use, both by aboriginal peoples and in modern culture.
- Among the Gnetales, Welwitschia has no known uses, but Ephedra has a long history of medicinal use, both by aboriginal peoples and in modern culture. Gnetum has produced some promising pharmacological compounds.
- The Pinales, Araucariales and Cupressales collectively comprise the conifers, and the use of this group by humans is legion. Numerous species in this group served aboriginal peoples for their wood, used for fuel, housing, boats, and tools. The resins were used as adhesives or sealants. Roots were commonly a source of fiber, as was bark sometimes. Seeds were a source of food, and the inner bark was also sometimes eaten or fed to animals. Organochemical extractives from every part of the plant found medicinal uses. Many species also had religious or spiritual significance; this was particularly true with yews (Taxus spp.) and with exceptionally large or ancient trees. Nearly all of these uses have continued into modern times, with commercial harvest of wood, bark, resins and seeds all happening to an economically significant extent, and a continued reverence for these trees represented in many programs to preserve and celebrate exceptional trees.
Today, of course, gymnosperms provide the world with softwood lumber and with most of the wood pulp supply. Most of this timber comes from a handful of species, notably Pinus radiata, the most widely planted tree in the world, although it has one of the smallest natural distributions.
Conifers are one of the world's most important renewable resources. Most economic and cultural exploitation concerns members of the families Pinaceae and Cupressaceae, with Araucariaceae (in Australia and South America) and Podocarpaceae (in scattered tropical locales) locally important.
Several conifers, particularly in the genera Agathis (Araucariaceae), Abies, and Pinus (Pinaceae), produce economically important resins. Resins are sticky, liquid, organic substances that usually harden when exposed to air into brittle, amorphous, solid substances. Natural resins are classified according to their physical and chemical properties into hard resins, oleoresins, and gum resins (Moussouris and Regato 1999).
Conifers have a long and complex spiritual tradition in our culture. Their evergreen nature has made them symbols of immortality; Taxus baccata is a good example. Spiritual powers were also attributed to the cedar of Lebanon, Cedrus libani. Other trees were so important to the lives of aboriginal peoples that they were personified or worshipped; examples include the western redcedar, Thuja plicata; the Bunya pine, Araucaria bidwillii; and the kauri, Agathis australis. Even in our modern secular culture, people still speak reverently about species such as the Coast redwood (Sequoia sempervirens) and the ahuehuete (Taxodium mucronatum).
Observations
Gymnosperm diversity is best observed at botanical gardens and arboreta. Many species of conifers, at least, are also well represented in ornamental collections throughout the temperate zones of the world.
Remarks
All trees, indeed all vascular plants, possess a fundamental architecture prescribed by their genetic code. This architecture remains constant throughout the plant's life, providing a uniform functional structure even as the plant experiences immense changes in size, from seed to tree, and even in spite of calamities that may befall the plant during its life. Every tree can be defined by one of 23 different architectural models. This is a complex topic, not yet fully explored, and I have not yet found a source that classifies the gymnosperms according to their various architectural models; for further information see this excellent blog and the additional sources cited therein.
Conifers reproduce by means of seeds borne on the scales of female cones, and the pollen is produced in separate male cones. Pollination in conifers is always dependent on wind currents to blow the abundant yellow pollen from the male cones to the female cones.
Citations
Ahuja, M. R., and D. B. Neale. 2005. Evolution of genome size in conifers. Silvae Genetica 54(3):126–137. doi:10.1515/sg-2005-0020.
Arrondo, O. G., and B. Petriella. 1980. Alicurá, nueva localidad plantífera Liásica, de la provincia de Neuquén, Argentina. Ameghiniana 17:200-215.
Bechem, E.E. and I.J. Alexander. 2012. Mycorrhiza status of Gnetum spp. in Cameroon: evaluating diversity with a view to ameliorating domestication efforts. Mycorrhiza 22(2):99-108.
Brundrett, Mark. 2008. Mycorrhizal Associations: The Web Resource. mycorrhizas.info, accessed 2009.06.09.
Castillo, G. R. H. 2001. Seed plant phylogeny and the anthophyte hypothesis. http://www.biology.ualberta.ca/courses.hp/biol606/OldLecs/Lecture2001.03.HC.html, accessed 2007.11.26, now defunct.
S.C. Collier, C.T. Yarnes, and R.P. Herman. 2003. Mycorrhizal dependency of Chihuahuan Desert plants is influenced by life history and root morphology. J. Arid Environ. 55:223-229.
Crane, P. R. 1988. Major clades and relationships in "higher" gymnosperms. Pp. 218-272 in C. B. Beck (ed.), Origin and Evolution of Gymnosperms. New York: Columbia University Press.
Crane, P. R. and G. R. Upchurch Jr. 1987. Drewria potomacensis gen. et sp. nov., an Early Cretaceous member of Gnetales from the Potomac Group of Virginia. American Journal of Botany 74:1723-1738.
Cronquist, A., A. Takhtajan, and W. Zimmermann. 1966. On the higher taxa of Embryophyta. Taxon 15:129-134.
[DEWHA] Department of the Environment, Water, Heritage and the Arts (2010). Lomatia tasmanica in Species Profile and Threats Database, Department of the Environment, Water, Heritage and the Arts, Canberra. www.environment.gov.au/sprat, accessed 2010.03.29.
Escapa, I. , R. Cúneo, and B. Axsmith. 2008. A new genus of the Cupressaceae (sensu lato) from the Jurassic of Patagonia: Implications for conifer megasporangiate cone homologies. Review of Palaeobotany and Palynology 151:110-122.
Fisher, Jack and Andrew Vovides. 2004. Mycorrhizae are present in cycad roots. The Botanical Review 70:16-23.
Fontana, Anna. 2006. Vesicular-arbuscular mycorrhizae of Ginkgo biloba L. in natural and controlled conditions. New Phytologist 99(3):441-447.
Gao, Z. and B. A. Thomas. 1989. A review of fossil cycad megasporophylls, with new evidence of Crossozamia pomel and its associated leaves from the lower Permian of Taiyuan, China. Review of Palaeobotany and Palynology 60(3):205-223.
Gorozh. 1904. Lekts. Morf. Sist. Archegon., pp. 72 and 88.
Hill, Ken. 2005. Diversity and evolution of gymnosperms. Pp. 25-44 in R. J. Henry, Plant Diversity and Evolution. Cambridge, MA: CABI Publishing.
Hilton, J. 2014. The origin of modern conifer families. www.birmingham.ac.uk/research/activity/geosystems/projects/conifer-families/index.aspx, accessed 2014.12.06.
IUCN. 2020. IUCN Red List version 2020-1: Table 4b: Red List Category summary for all plant classes and families. https://nc.iucnredlist.org/redlist/content/attachment_files/2020_1_RL_Stats_Table_4b.pdf, accessed 2022.09.23.
K.M. Jacobson, P.J. Jacobson, and O.K. Miller Jr. 1993. The mycorrhizal status of Welwitschia mirabilis. Mycorrhiza 3(1):13-17.
Leslie, A. B., J. Beaulieu, G. Holman, C. S. Campbell, W. Mei, L. R. Raubeson, and S. Mathews. 2018. An overview of extant conifer evolution from the perspective of the fossil record. American Journal of Botany 105(9):1531–1544.
Li, Z., Baniaga, A. E., Sessa, E. B., Scascitelli, M., Graham, S. W., Rieseberg, L. H., and Barker, M. S. 2015. Early genome duplications in conifers and other seed plants. Science Advances 1(10):e1501084–e1501084. doi:10.1126/sciadv.1501084.
Link, J. H. F. 1829. Handbuch zur Erkennung der nutzbarsten und am haüfigsten vorkommenden Gewächse. Berlin. V. 2, p. 470.
Liu, Y., et al. 2022. The Cycas genome and the early evolution of seed plants. Nat. Plants 1–13. Nature Publishing Group. doi:10.1038/s41477-022-01129-7.
Nathorst, A. G. 1908. Paläobotanische Mitteilungen, 7: Über Palissya, Stachyotaxus und Palaeotaxus. Kungliga Svenska Vetenskapsakademiens
Handlingar 43:3-37.
Ohsawa, T., M. Nishida, and H. Nishida. 1991. Structure and affinities of the petrified plants from the Cretaceous of northern Japan and Saghalien IX. A petrified cone of Sciadopitys from the Upper Cretaceous of Hokkaido. Journal of Phytogeography and Taxonomy 39:97-105.
Ran, J.-H., T.-T. Shen, M.-M. Wang, and X.-Q. Wang. 2018. Phylogenomics resolves the deep phylogeny of seed plants and indicates partial convergent or homoplastic evolution between Gnetales and angiosperms. Proc. R. Soc. B. 285(1881):20181012.
Rothwell, G. W., G. Mapes, R. A. Stockey, and J. Hilton. 2012. The seed cone Eathiestrobus gen. nov.: fossil evidence for a Jurassic origin of Pinaceae. American Journal of Botany 99(4):708-720.
Ryberg, P. E., G. W. Rothwell, R. A. Stockey, J. Hilton, G. Mapes, and J. B. Riding. 2012. Reconsidering relationships among stem and crown group Pinaceae: oldest record of the genus Pinus from the Early Cretaceous of Yorkshire, United Kingdom. International Journal of Plant Sciences 173(8):917-932.
Tedersoo, L. and S. Põlme. 2012. Infrageneric variation in partner specificity: multiple ectomycorrhizal symbionts associate with Gnetum gnemon (Gnetophyta) in Papua New Guinea. Mycorrhiza 22(8):663-8.
Thomas, B. A., and Spicer, R. A. 1986. The Evolution and Palaeobiology of Land Plants. London: Croom Helm. ix, 309p.
Tomlinson, P. B. 1983. Tree Architecture: New approaches help to define the elusive biological property of tree form. American Scientist 71(2):141-149. Available http://harvardforest.fas.harvard.edu/sites/harvardforest.fas.harvard.edu/files/publications/pdfs/Tomlinson_AmScientist_1983.pdf, accessed 2017.08.12.
Townrow, J. A. 1967. On Rissikia and Mataia - Podocarpaceous conifers
from the lower Mesozoic of southern lands. Papers and Proceedings of the Royal Society of Tasmania 101:103-136.
Tralau, H. 1968. Evolutionary trends in the genus Ginkgo. Lethaia 1:63–101.
Walas, Łukasz., Wojciech Mandryk, Peter A. Thomas, Zanna Tyrała-Wierucka, and Grzegorz Iszkuło. 2018. Sexual systems in gymnosperms: a review. Basic and Applied Ecology 31:1-9. https://doi.org/10.1016/j.baae.2018.05.009.
Wang, X.-Q., and J.-H. Ran. 2014. Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution 75:24–40.
See also
Burleigh, J. G. and S. Mathews. 2004. Phylogenetic signal in nucleotide data from seed plants: implications for resolving the seed plant tree of life. American Journal of Botany 91: 1599-1613. Available online, accessed 2007.11.26.
Chaw, S.M., C.L. Parkinson, Y. Cheng, T.M. Vincent, and J.D. Palmer. 2000. Seed plant phylogeny inferred frrom all three plant genomes:
monophyly of extant gymnosperms and origin of Gnetales from conifers. Proc Natl Acad Sci USA 97:4086-4091. Available online HERE (2007.11.26).
Christenhusz, M. J. M., J. L. Reveal, A. Farjon, M. F. Gardner, R. R. Mill, and M. W. Chase. 2011. A new classification and linear sequence of extant gymnosperms. Phytotaxa 19:55-70.
Eckenwalder 2009.
Enright and Hill 1990.
Farjon 2010.
The University of Sydney School of Biological Sciences provides a good overview of the gymnosperm life cycle (2006.03.20).
Wang, Xiao-Quan, and Jin-Hua Ran. 2014. Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution 75:24-40.
White 1994 gives a beautifully illustrated account of the fossil origins of the major gymnosperm and angiosperm groups.